Lan Fei, Collins Robert E, De Cegli Rossella, Alpatov Roman, Horton John R, Shi Xiaobing, Gozani Or, Cheng Xiaodong, Shi Yang
Department of Pathology, Harvard Medical School, 77 Ave Louis Pasteur, Boston, Massachusetts 02115, USA.
Nature. 2007 Aug 9;448(7154):718-22. doi: 10.1038/nature06034.
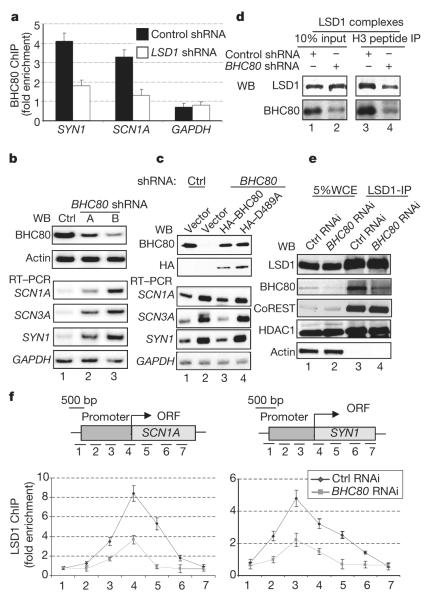
Histone methylation is crucial for regulating chromatin structure, gene transcription and the epigenetic state of the cell. LSD1 is a lysine-specific histone demethylase that represses transcription by demethylating histone H3 on lysine 4 (ref. 1). The LSD1 complex contains a number of proteins, all of which have been assigned roles in events upstream of LSD1-mediated demethylation apart from BHC80 (also known as PHF21A), a plant homeodomain (PHD) finger-containing protein. Here we report that, in contrast to the PHD fingers of the bromodomain PHD finger transcription factor (BPTF) and inhibitor of growth family 2 (ING2), which bind methylated H3K4 (H3K4me3), the PHD finger of BHC80 binds unmethylated H3K4 (H3K4me0), and this interaction is specifically abrogated by methylation of H3K4. The crystal structure of the PHD finger of BHC80 bound to an unmodified H3 peptide has revealed the structural basis of the recognition of H3K4me0. Knockdown of BHC80 by RNA inhibition results in the de-repression of LSD1 target genes, and this repression is restored by the reintroduction of wild-type BHC80 but not by a PHD-finger mutant that cannot bind H3. Chromatin immunoprecipitation showed that BHC80 and LSD1 depend reciprocally on one another to associate with chromatin. These findings couple the function of BHC80 to that of LSD1, and indicate that unmodified H3K4 is part of the 'histone code'. They further raise the possibility that the generation and recognition of the unmodified state on histone tails in general might be just as crucial as post-translational modifications of histone for chromatin and transcriptional regulation.
组蛋白甲基化对于调节染色质结构、基因转录及细胞的表观遗传状态至关重要。LSD1是一种赖氨酸特异性组蛋白去甲基化酶,可通过使组蛋白H3赖氨酸4位点去甲基化来抑制转录(参考文献1)。LSD1复合物包含多种蛋白质,除了BHC80(也称为PHF21A,一种含植物同源结构域(PHD)指的蛋白质)外,所有这些蛋白质在LSD1介导的去甲基化上游事件中都发挥了作用。我们在此报告,与结合甲基化H3K4(H3K4me3)的溴结构域PHD指转录因子(BPTF)和生长抑制家族2(ING2)的PHD指不同,BHC80的PHD指结合未甲基化的H3K4(H3K4me0),并且这种相互作用会被H3K4的甲基化特异性消除。BHC80的PHD指与未修饰的H3肽结合的晶体结构揭示了对H3K4me0识别的结构基础。通过RNA干扰敲低BHC80会导致LSD1靶基因的去抑制,并且通过重新引入野生型BHC80可恢复这种抑制,但不能通过不能结合H3的PHD指突变体恢复。染色质免疫沉淀显示,BHC80和LSD1相互依赖才能与染色质结合。这些发现将BHC80的功能与LSD1的功能联系起来,并表明未修饰的H3K4是“组蛋白密码”的一部分。它们进一步增加了一种可能性,即一般来说,组蛋白尾部未修饰状态的产生和识别对于染色质和转录调控可能与组蛋白的翻译后修饰同样重要。