Thomas Sean, Green Amanda, Sturm Nancy R, Campbell David A, Myler Peter J
Department of Genome Sciences, University of Washington, Seattle, WA 98195, USA.
BMC Genomics. 2009 Apr 8;10:152. doi: 10.1186/1471-2164-10-152.
Many components of the RNA polymerase II transcription machinery have been identified in kinetoplastid protozoa, but they diverge substantially from other eukaryotes. Furthermore, protein-coding genes in these organisms lack individual transcriptional regulation, since they are transcribed as long polycistronic units. The transcription initiation sites are assumed to lie within the 'divergent strand-switch' regions at the junction between opposing polycistronic gene clusters. However, the mechanism by which Kinetoplastidae initiate transcription is unclear, and promoter sequences are undefined.
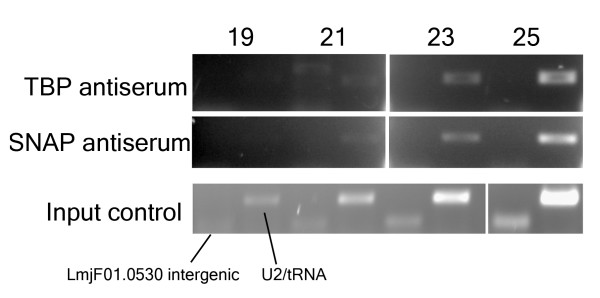
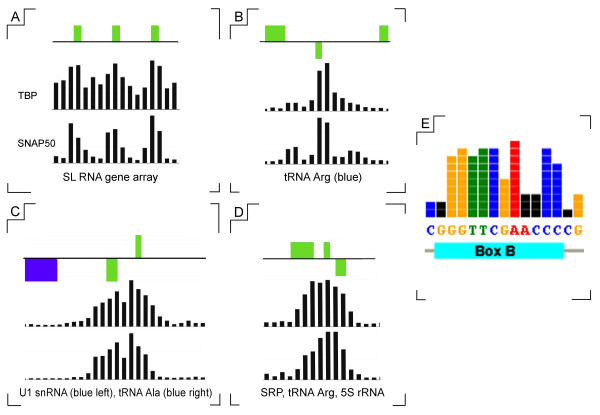
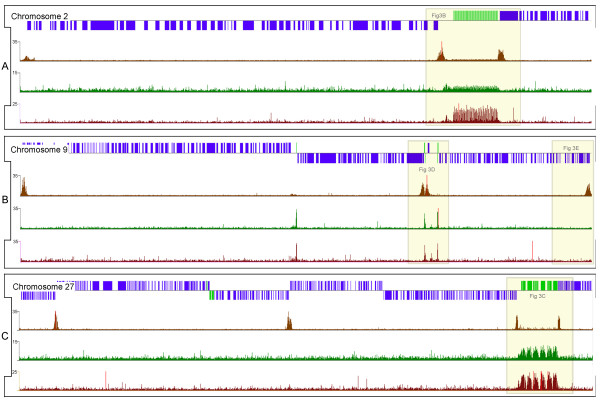
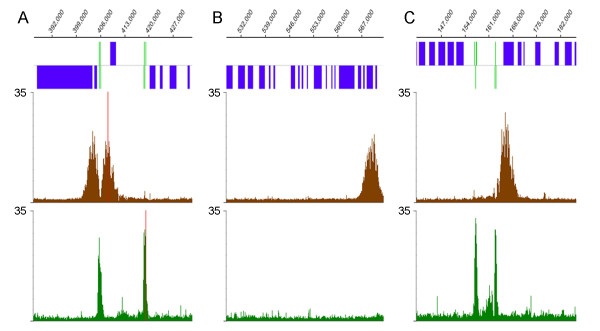
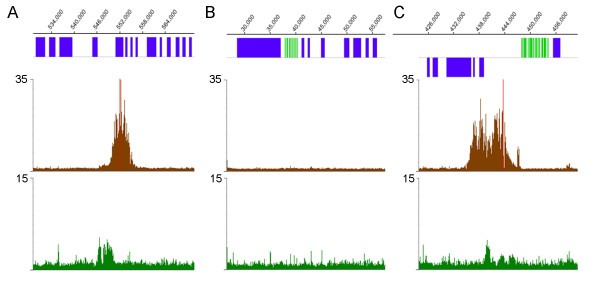
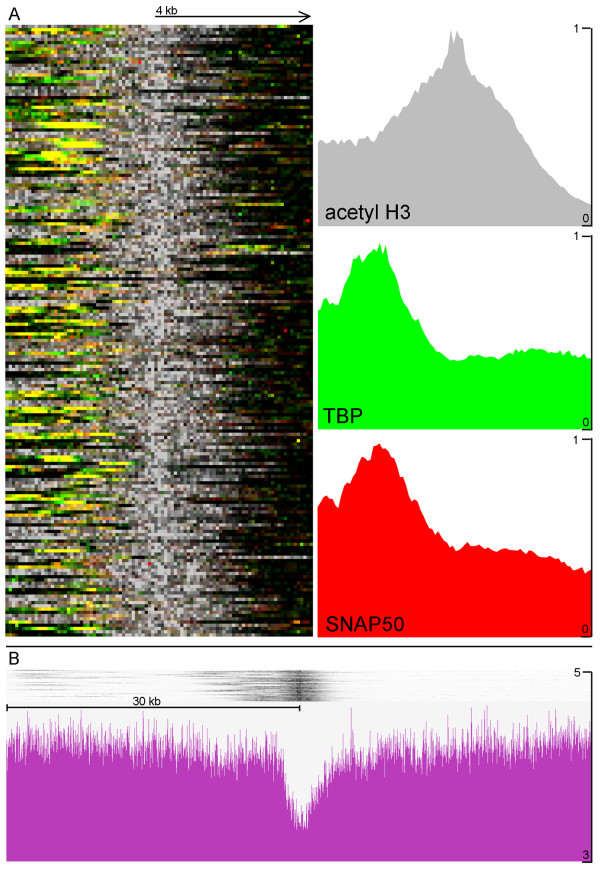
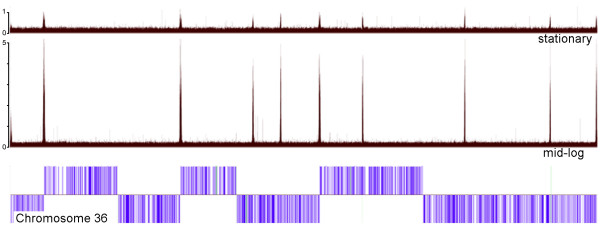
The chromosomal location of TATA-binding protein (TBP or TRF4), Small Nuclear Activating Protein complex (SNAP50), and H3 histones were assessed in Leishmania major using microarrays hybridized with DNA obtained through chromatin immunoprecipitation (ChIP-chip). The TBP and SNAP50 binding patterns were almost identical and high intensity peaks were associated with tRNAs and snRNAs. Only 184 peaks of acetylated H3 histone were found in the entire genome, with substantially higher intensity in rapidly-dividing cells than stationary-phase. The majority of the acetylated H3 peaks were found at divergent strand-switch regions, but some occurred at chromosome ends and within polycistronic gene clusters. Almost all these peaks were associated with lower intensity peaks of TBP/SNAP50 binding a few kilobases upstream, evidence that they represent transcription initiation sites.
The first genome-wide maps of DNA-binding protein occupancy in a kinetoplastid organism suggest that H3 histones at the origins of polycistronic transcription of protein-coding genes are acetylated. Global regulation of transcription initiation may be achieved by modifying the acetylation state of these origins.
在动基体原生动物中已鉴定出RNA聚合酶II转录机制的许多成分,但它们与其他真核生物有很大差异。此外,这些生物体中的蛋白质编码基因缺乏个体转录调控,因为它们作为长的多顺反子单元进行转录。转录起始位点被认为位于相对的多顺反子基因簇之间连接处的“发散链切换”区域内。然而,锥虫科启动转录的机制尚不清楚,启动子序列也未明确。
使用与通过染色质免疫沉淀(ChIP-chip)获得的DNA杂交的微阵列,在硕大利什曼原虫中评估了TATA结合蛋白(TBP或TRF4)、小核激活蛋白复合物(SNAP50)和H3组蛋白的染色体定位。TBP和SNAP50的结合模式几乎相同,高强度峰与tRNA和snRNA相关。在整个基因组中仅发现184个乙酰化H3组蛋白峰,在快速分裂的细胞中强度明显高于静止期。大多数乙酰化H3峰位于发散链切换区域,但有些出现在染色体末端和多顺反子基因簇内。几乎所有这些峰都与上游几千碱基处TBP/SNAP50结合的较低强度峰相关,这证明它们代表转录起始位点。
动基体生物中首个全基因组DNA结合蛋白占据图谱表明,蛋白质编码基因多顺反子转录起始处的H3组蛋白被乙酰化。转录起始的全局调控可能通过改变这些起始位点的乙酰化状态来实现。