MRC Protein Phosphorylation Unit, College of Life Sciences, University of Dundee, Dundee DD15EH, Scotland, U.K.
Biochem J. 2010 Mar 15;427(1):69-78. doi: 10.1042/BJ20091834.
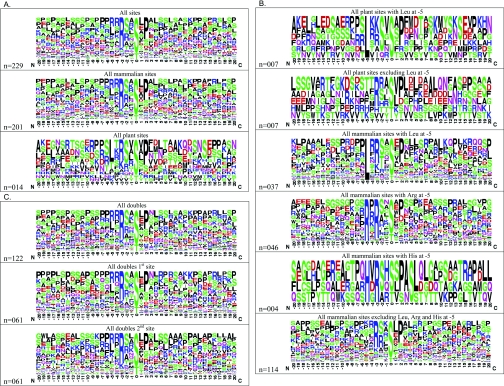
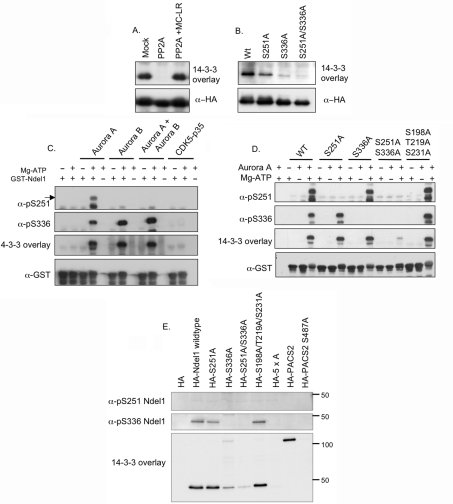
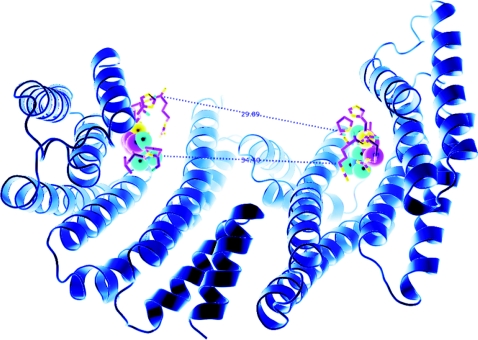
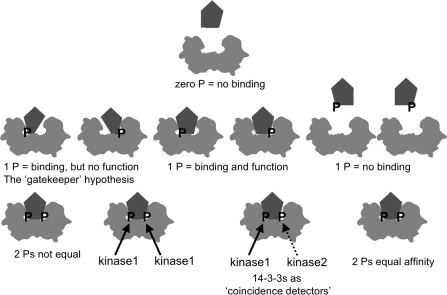
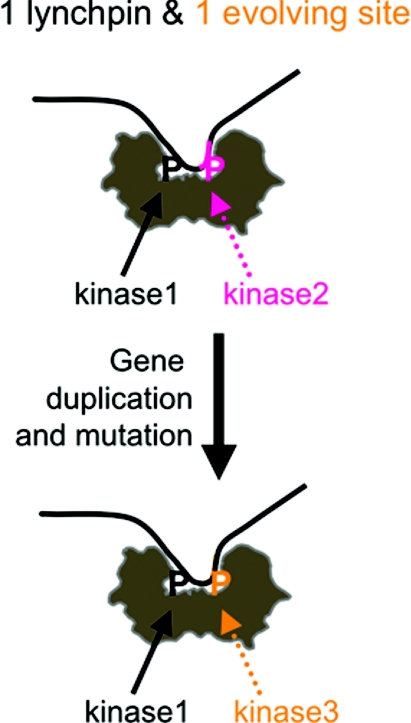
More than 200 phosphorylated 14-3-3-binding sites in the literature were analysed to define 14-3-3 specificities, identify relevant protein kinases, and give insights into how cellular 14-3-3/phosphoprotein networks work. Mode I RXX(pS/pT)XP motifs dominate, although the +2 proline residue occurs in less than half, and LX(R/K)SX(pS/pT)XP is prominent in plant 14-3-3-binding sites. Proline at +1 is rarely reported, and such motifs did not stand up to experimental reanalysis of human Ndel1. Instead, we discovered that 14-3-3 interacts with two residues that are phosphorylated by basophilic kinases and located in the DISC1 (disrupted-in-schizophrenia 1)-interacting region of Ndel1 that is implicated in cognitive disorders. These data conform with the general findings that there are different subtypes of 14-3-3-binding sites that overlap with the specificities of different basophilic AGC (protein kinase A/protein kinase G/protein kinase C family) and CaMK (Ca2+/calmodulin-dependent protein kinase) protein kinases, and a 14-3-3 dimer often engages with two tandem phosphorylated sites, which is a configuration with special signalling, mechanical and evolutionary properties. Thus 14-3-3 dimers can be digital logic gates that integrate more than one input to generate an action, and coincidence detectors when the two binding sites are phosphorylated by different protein kinases. Paired sites are generally located within disordered regions and/or straddle either side of functional domains, indicating how 14-3-3 dimers modulate the conformations and/or interactions of their targets. Finally, 14-3-3 proteins bind to members of several multi-protein families. Two 14-3-3-binding sites are conserved across the class IIa histone deacetylases, whereas other protein families display differential regulation by 14-3-3s. We speculate that 14-3-3 dimers may have contributed to the evolution of such families, tailoring regulatory inputs to different physiological demands.
对文献中超过 200 个磷酸化的 14-3-3 结合位点进行了分析,以确定 14-3-3 的特异性,确定相关的蛋白激酶,并深入了解细胞内 14-3-3/磷酸蛋白网络的工作方式。模式 I 的 RXX(pS/pT)XP 基序占主导地位,尽管 +2 脯氨酸残基不到一半,而植物 14-3-3 结合位点中 LX(R/K)SX(pS/pT)XP 则很突出。+1 位脯氨酸很少有报道,而且这些基序在对人类 Ndel1 的实验重新分析中也站不住脚。相反,我们发现 14-3-3 与两个由碱性激酶磷酸化的残基相互作用,这些残基位于 Ndel1 的 DISC1(精神分裂症 1 中缺失)相互作用区域,该区域与认知障碍有关。这些数据与一般发现一致,即存在不同类型的 14-3-3 结合位点,这些结合位点与不同的碱性 AGC(蛋白激酶 A/蛋白激酶 G/蛋白激酶 C 家族)和 CaMK(Ca2+/钙调蛋白依赖性蛋白激酶)蛋白激酶的特异性重叠,并且 14-3-3 二聚体通常与两个串联磷酸化位点结合,这是一种具有特殊信号、机械和进化特性的构型。因此,14-3-3 二聚体可以是数字逻辑门,它可以整合多个输入来产生一个动作,并且当两个结合位点被不同的蛋白激酶磷酸化时,它可以作为一个巧合检测器。配对的位点通常位于无序区域内和/或跨越功能域的两侧,这表明 14-3-3 二聚体如何调节其靶标的构象和/或相互作用。最后,14-3-3 蛋白与几个多蛋白家族的成员结合。两个 14-3-3 结合位点在 IIa 类组蛋白去乙酰化酶中是保守的,而其他蛋白家族则受到 14-3-3 的不同调节。我们推测,14-3-3 二聚体可能有助于这些家族的进化,根据不同的生理需求调整调节输入。