Department of Microbiology, University of California, Davis, Davis, California 95616-8665, USA.
Nature. 2010 Sep 2;467(7311):112-6. doi: 10.1038/nature09355.
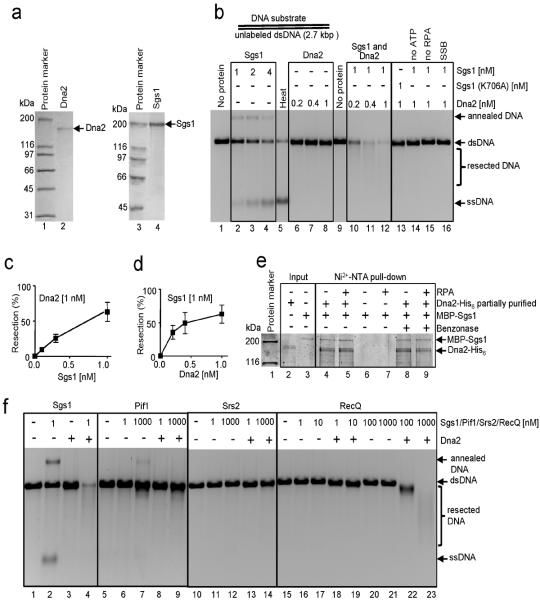
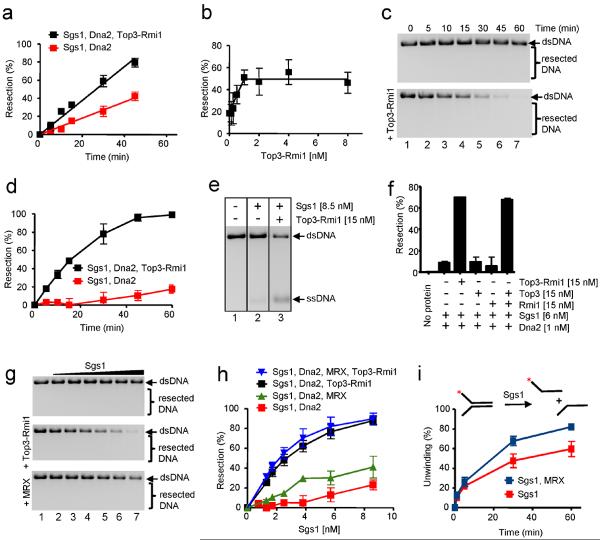
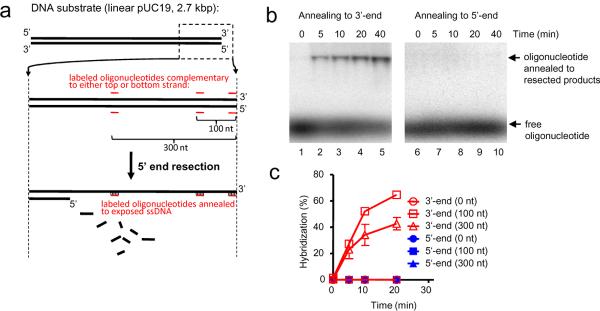
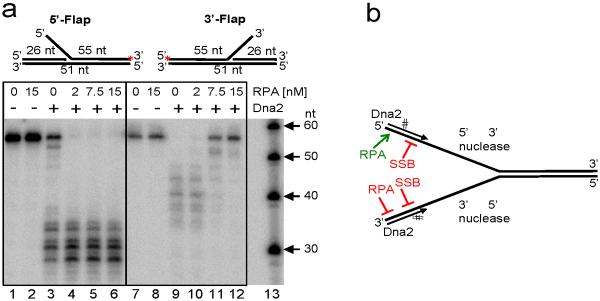
The repair of DNA double-strand breaks (DSBs) by homologous recombination requires processing of broken ends. For repair to start, the DSB must first be resected to generate a 3'-single-stranded DNA (ssDNA) overhang, which becomes a substrate for the DNA strand exchange protein, Rad51 (ref. 1). Genetic studies have implicated a multitude of proteins in the process, including helicases, nucleases and topoisomerases. Here we biochemically reconstitute elements of the resection process and reveal that it requires the nuclease Dna2, the RecQ-family helicase Sgs1 and the ssDNA-binding protein replication protein-A (RPA). We establish that Dna2, Sgs1 and RPA constitute a minimal protein complex capable of DNA resection in vitro. Sgs1 helicase unwinds the DNA to produce an intermediate that is digested by Dna2, and RPA stimulates DNA unwinding by Sgs1 in a species-specific manner. Interestingly, RPA is also required both to direct Dna2 nucleolytic activity to the 5'-terminated strand of the DNA break and to inhibit 3' to 5' degradation by Dna2, actions that generate and protect the 3'-ssDNA overhang, respectively. In addition to this core machinery, we establish that both the topoisomerase 3 (Top3) and Rmi1 complex and the Mre11-Rad50-Xrs2 complex (MRX) have important roles as stimulatory components. Stimulation of end resection by the Top3-Rmi1 heterodimer and the MRX proteins is by complex formation with Sgs1 (refs 5, 6), which unexpectedly stimulates DNA unwinding. We suggest that Top3-Rmi1 and MRX are important for recruitment of the Sgs1-Dna2 complex to DSBs. Our experiments provide a mechanistic framework for understanding the initial steps of recombinational DNA repair in eukaryotes.
DNA 双链断裂 (DSBs) 的同源重组修复需要对断裂末端进行加工。为了启动修复,DSB 必须首先被切除以产生 3'-单链 DNA (ssDNA) 突出端,该突出端成为 DNA 链交换蛋白 Rad51 的底物(参考文献 1)。遗传研究表明,许多蛋白质参与了这个过程,包括解旋酶、核酸酶和拓扑异构酶。在这里,我们通过生化方法重新构建了切除过程的元件,并揭示它需要核酸酶 Dna2、RecQ 家族解旋酶 Sgs1 和 ssDNA 结合蛋白复制蛋白-A (RPA)。我们确定 Dna2、Sgs1 和 RPA 构成了一个能够在体外进行 DNA 切除的最小蛋白复合物。Sgs1 解旋酶解开 DNA 产生一个中间产物,该产物被 Dna2 消化,RPA 以种属特异性的方式刺激 Sgs1 的 DNA 解旋。有趣的是,RPA 还需要将 Dna2 核酶活性引导到 DNA 断裂的 5'-端,并抑制 Dna2 的 3'-5' 降解,这两个作用分别产生和保护 3'-ssDNA 突出端。除了这个核心机制外,我们还确定拓扑异构酶 3 (Top3) 和 Rmi1 复合物以及 Mre11-Rad50-Xrs2 复合物 (MRX) 作为刺激因子具有重要作用。Top3-Rmi1 异源二聚体和 MRX 蛋白对末端切除的刺激是通过与 Sgs1 形成复合物(参考文献 5,6),这出乎意料地刺激了 DNA 解旋。我们认为 Top3-Rmi1 和 MRX 对于将 Sgs1-Dna2 复合物招募到 DSB 至关重要。我们的实验为理解真核生物重组 DNA 修复的初始步骤提供了一个机制框架。