Departamento de Bioquímica, Biología Celular y Molecular de Plantas, Estación Experimental del Zaidín CSIC, Profesor Albareda 1, 18008 Granada, Spain.
J Exp Bot. 2011 Jan;62(3):1255-69. doi: 10.1093/jxb/erq354. Epub 2010 Nov 16.
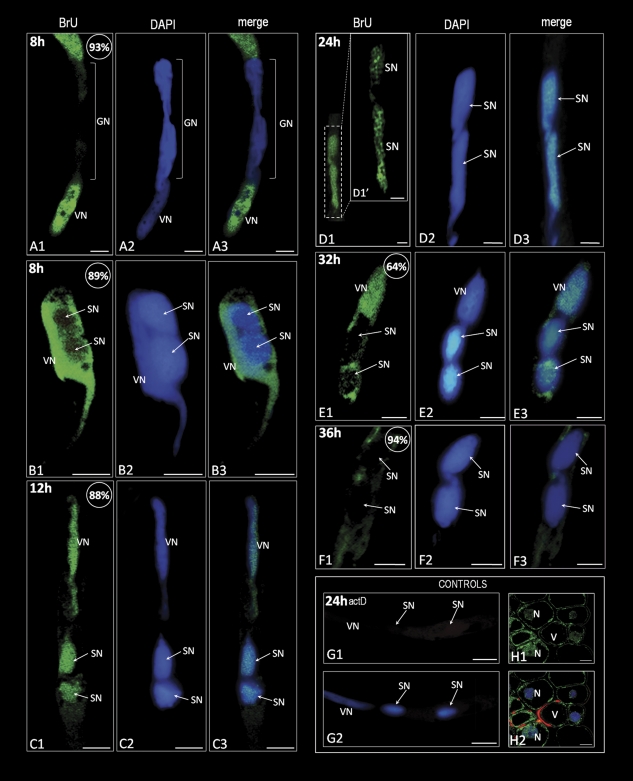
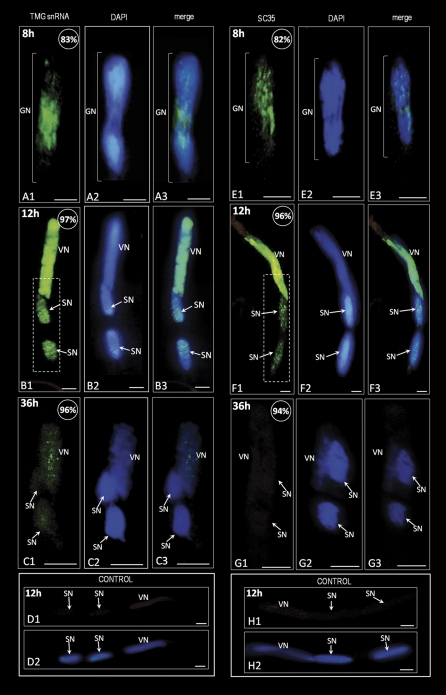
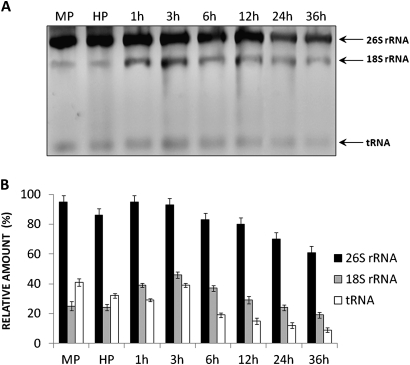
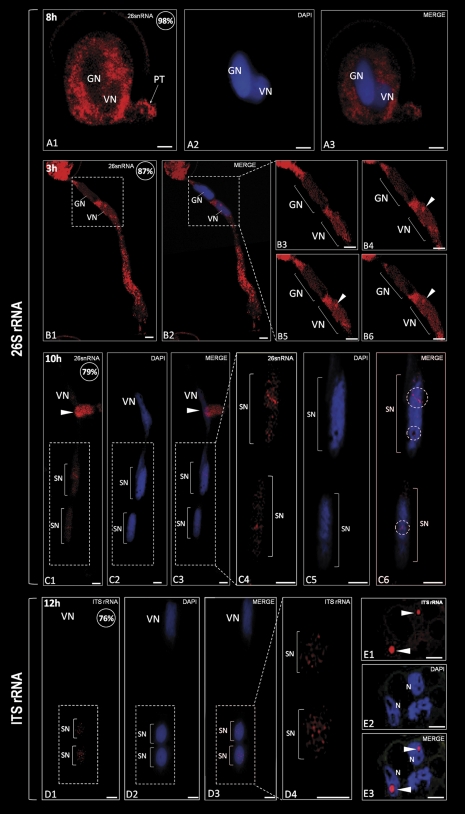
In this study, the transcriptional state and distribution of RNA polymerase II, pre-mRNA splicing machinery elements, and rRNA transcripts were investigated in the sperm cells of Hyacinthus orientalis L. during in vitro pollen tube growth. During the second pollen mitosis, no nascent transcripts were observed in the area of the dividing generative cell, whereas the splicing factors were present and their pools were divided between newly formed sperm cells. Just after their origin, the sperm cells were shown to synthesize new RNA, although at a markedly lower level than the vegetative nucleus. The occurrence of RNA synthesis was accompanied by the presence of RNA polymerase II and a rich pool of splicing machinery elements. Differences in the spatial pattern of pre-mRNA splicing factors localization reflect different levels of RNA synthesis in the vegetative nucleus and sperm nuclei. In the vegetative nucleus, they were localized homogenously, whereas in the sperm nuclei a mainly speckled pattern of small nuclear RNA with a trimethylguanosine cap (TMG snRNA) and SC35 protein distribution was observed. As pollen tube growth proceeded, inhibition of RNA synthesis in the sperm nuclei was observed, which was accompanied by a gradual elimination of the splicing factors. In addition, analysis of rRNA localization indicated that the sperm nuclei are likely to synthesize some pool of rRNA at the later steps of pollen tube. It is proposed that the described changes in the nuclear activity of H. orientalis sperm cells reflect their maturation process during pollen tube growth, and that mature sperm cells do not carry into the zygote the nascent transcripts or the splicing machinery elements.
在这项研究中,研究了东方风信子精子细胞中 RNA 聚合酶 II、前体 mRNA 剪接机制元件和 rRNA 转录本的转录状态和分布,这些精子细胞在体外花粉管生长过程中。在第二次花粉有丝分裂中,在分裂的生殖细胞区域没有观察到新生转录本,而剪接因子存在,并且它们的池在新形成的精子细胞之间分配。就在它们起源之后,精子细胞被证明可以合成新的 RNA,尽管其水平明显低于营养核。RNA 合成的发生伴随着 RNA 聚合酶 II 和丰富的剪接机制元件池的存在。前体 mRNA 剪接因子定位的空间模式差异反映了营养核和精子核中 RNA 合成水平的不同。在营养核中,它们均匀分布,而在精子核中,观察到富含小核 RNA(TMG snRNA)和 SC35 蛋白的斑点模式。随着花粉管生长,观察到精子核中 RNA 合成的抑制,这伴随着剪接因子的逐渐消除。此外,对 rRNA 定位的分析表明,精子核可能在花粉管生长的后期合成一些 rRNA 池。提出描述的东方风信子精子细胞核活性的变化反映了它们在花粉管生长过程中的成熟过程,并且成熟的精子细胞不会将新生转录本或剪接机制元件带入受精卵。