Department of Plant Biology, College of Biological Sciences, University of California Davis, Davis, California, United States of America.
PLoS Genet. 2011 Mar;7(3):e1001350. doi: 10.1371/journal.pgen.1001350. Epub 2011 Mar 31.
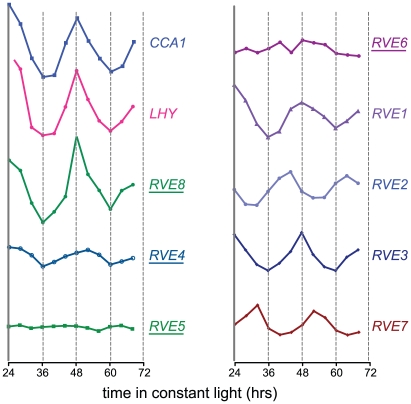
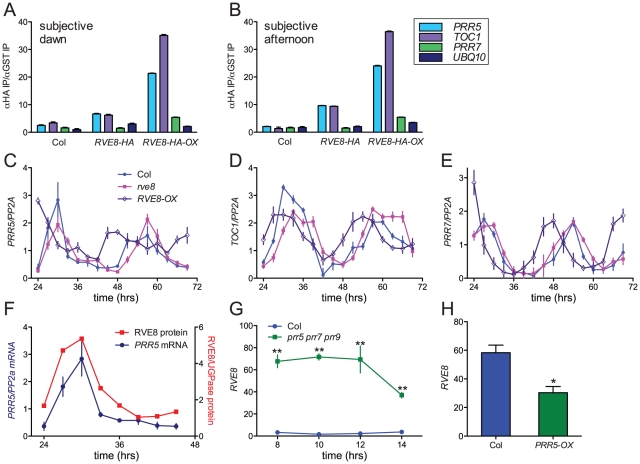
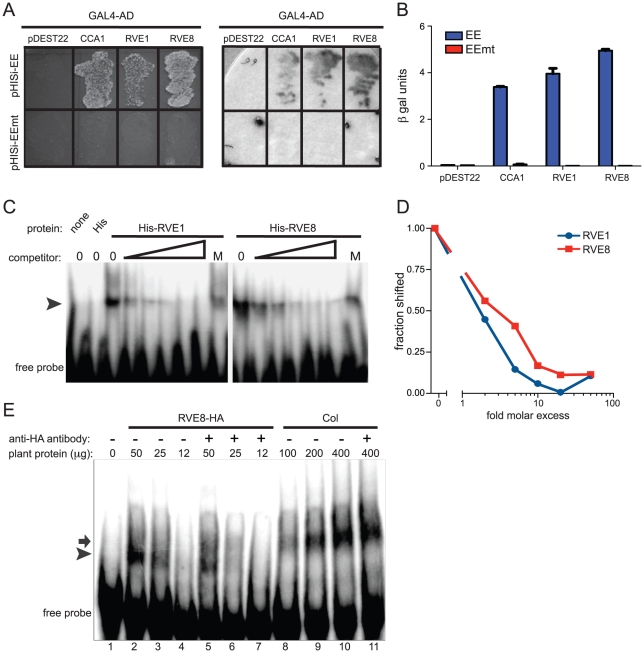
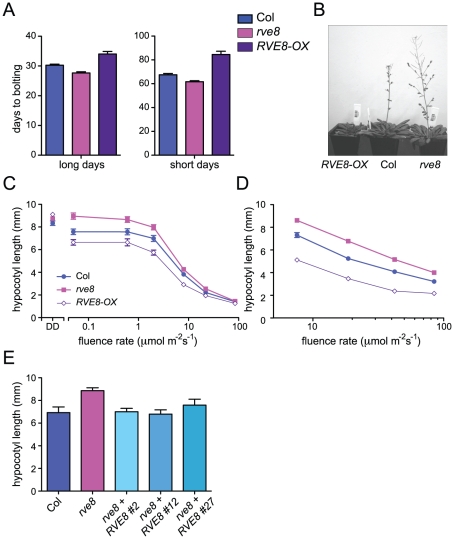
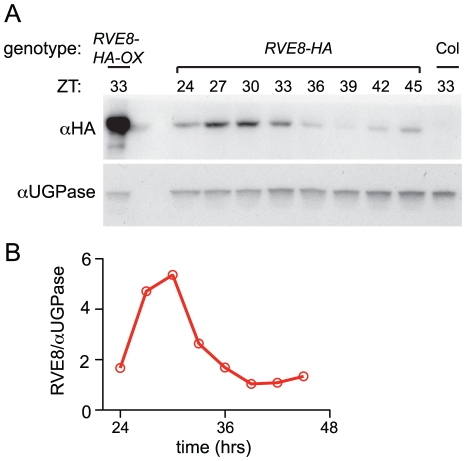
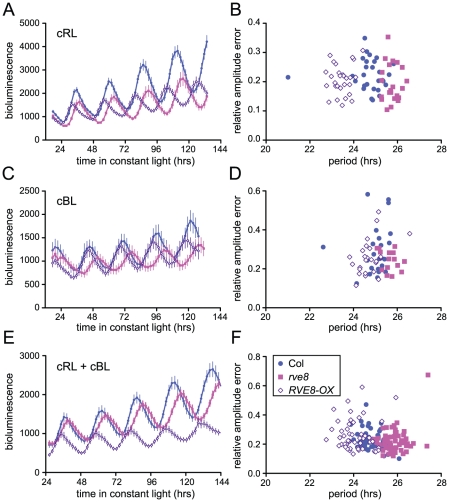
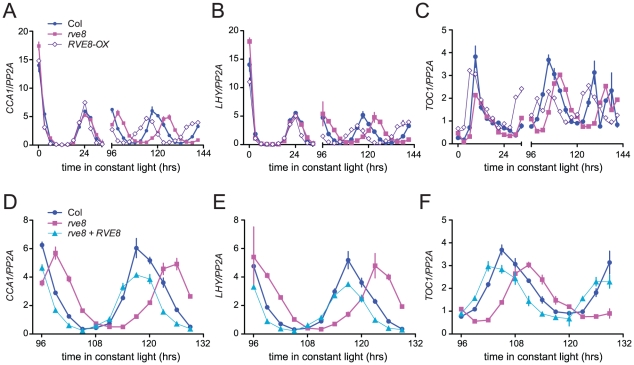
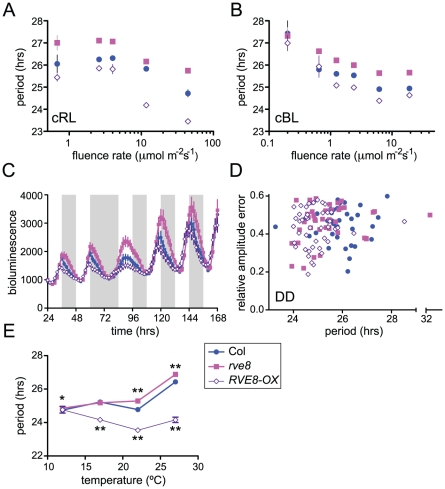
Circadian rhythms provide organisms with an adaptive advantage, allowing them to regulate physiological and developmental events so that they occur at the most appropriate time of day. In plants, as in other eukaryotes, multiple transcriptional feedback loops are central to clock function. In one such feedback loop, the Myb-like transcription factors CCA1 and LHY directly repress expression of the pseudoresponse regulator TOC1 by binding to an evening element (EE) in the TOC1 promoter. Another key regulatory circuit involves CCA1 and LHY and the TOC1 homologs PRR5, PRR7, and PRR9. Purification of EE-binding proteins from plant extracts followed by mass spectrometry led to the identification of RVE8, a homolog of CCA1 and LHY. Similar to these well-known clock genes, expression of RVE8 is circadian-regulated with a dawn phase of expression, and RVE8 binds specifically to the EE. However, whereas cca1 and lhy mutants have short period phenotypes and overexpression of either gene causes arrhythmia, rve8 mutants have long-period and RVE8-OX plants have short-period phenotypes. Light input to the clock is normal in rve8, but temperature compensation (a hallmark of circadian rhythms) is perturbed. RVE8 binds to the promoters of both TOC1 and PRR5 in the subjective afternoon, but surprisingly only PRR5 expression is perturbed by overexpression of RVE8. Together, our data indicate that RVE8 promotes expression of a subset of EE-containing clock genes towards the end of the subjective day and forms a negative feedback loop with PRR5. Thus RVE8 and its homologs CCA1 and LHY function close to the circadian oscillator but act via distinct molecular mechanisms.
昼夜节律为生物体提供了一种适应优势,使它们能够调节生理和发育事件,以便在一天中最合适的时间发生。在植物中,与其他真核生物一样,多个转录反馈回路是时钟功能的核心。在这样的一个反馈回路中,Myb 样转录因子 CCA1 和 LHY 通过结合 TOC1 启动子中的一个夜间元件(EE),直接抑制拟应答调节因子 TOC1 的表达。另一个关键的调节回路涉及 CCA1 和 LHY 以及 TOC1 同源物 PRR5、PRR7 和 PRR9。从植物提取物中纯化 EE 结合蛋白,然后进行质谱分析,导致鉴定出 RVE8,它是 CCA1 和 LHY 的同源物。与这些著名的生物钟基因相似,RVE8 的表达具有昼夜节律调节的特征,表现出黎明阶段的表达,并且 RVE8 特异性地结合 EE。然而,cca1 和 lhy 突变体具有短周期表型,并且这两个基因的过表达导致节律失常,而 rve8 突变体具有长周期,并且 RVE8-OX 植物具有短周期表型。RVE8 对时钟的光输入是正常的,但温度补偿(昼夜节律的一个标志)受到干扰。RVE8 在主观下午与 TOC1 和 PRR5 的启动子结合,但令人惊讶的是,只有 PRR5 的表达受到 RVE8 过表达的干扰。我们的数据表明,RVE8 促进了一组含有 EE 的时钟基因在主观日接近尾声时的表达,并与 PRR5 形成了一个负反馈回路。因此,RVE8 及其同源物 CCA1 和 LHY 靠近生物钟振荡器起作用,但通过不同的分子机制起作用。