Institute of Biological, Environmental and Rural Sciences, Aberystwyth University, Penglais, Aberystwyth, SY23 3DA, UK.
BMC Evol Biol. 2011 Aug 10;11:236. doi: 10.1186/1471-2148-11-236.
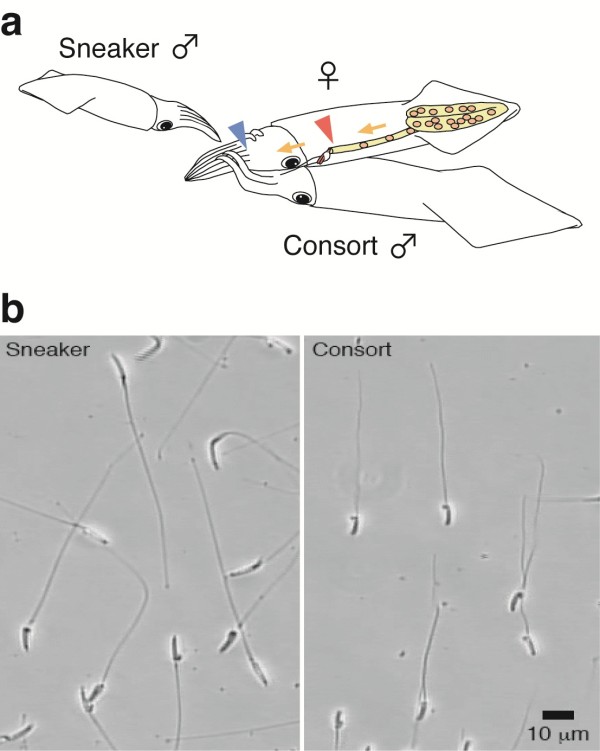
Sperm cells are the target of strong sexual selection that may drive changes in sperm structure and function to maximize fertilisation success. Sperm evolution is regarded to be one of the major consequences of sperm competition in polyandrous species, however it can also be driven by adaptation to the environmental conditions at the site of fertilization. Strong stabilizing selection limits intra-specific variation, and therefore polymorphism, among fertile sperm (eusperm). Here we analyzed reproductive morphology differences among males employing characteristic alternative mating behaviours, and so potentially different conditions of sperm competition and fertilization environment, in the squid Loligo bleekeri.
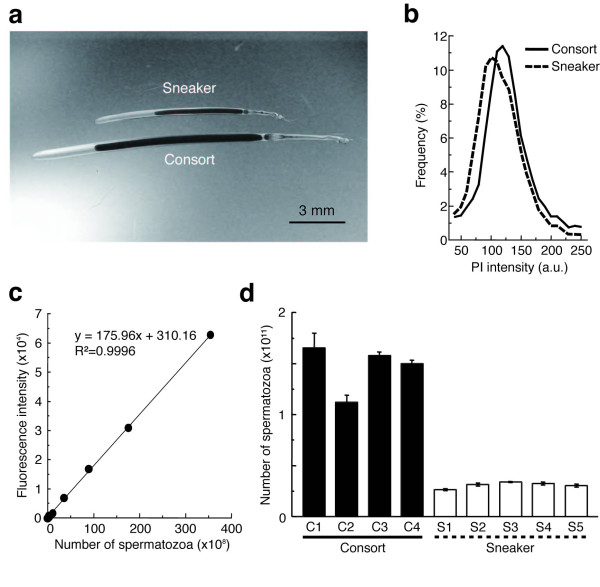
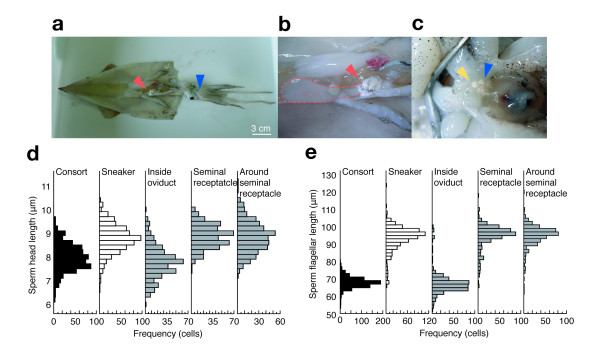
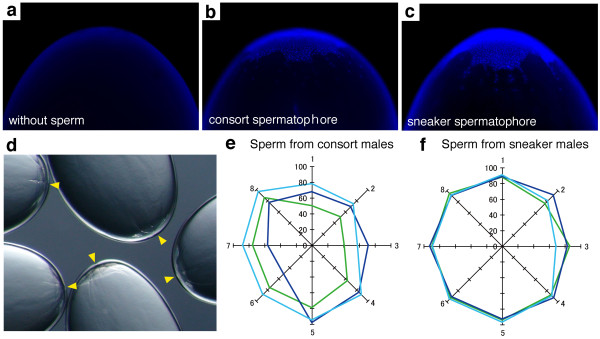
Large consort males transfer smaller (average total length = 73 μm) sperm to a female's internal sperm storage location, inside the oviduct; whereas small sneaker males transfer larger (99 μm) sperm to an external location around the seminal receptacle near the mouth. No significant difference in swimming speed was observed between consort and sneaker sperm. Furthermore, sperm precedence in the seminal receptacle was not biased toward longer sperm, suggesting no evidence for large sperm being favoured in competition for space in the sperm storage organ among sneaker males.
Here we report the first case, in the squid Loligo bleekeri, where distinctly dimorphic eusperm are produced by different sized males that employ alternative mating behaviours. Our results found no evidence that the distinct sperm dimorphism was driven by between- and within-tactic sperm competition. We propose that presence of alternative fertilization environments with distinct characteristics (i.e. internal or external), whether or not in combination with the effects of sperm competition, can drive the disruptive evolution of sperm size.
精子是强烈的性选择的目标,这可能导致精子结构和功能的变化,以最大限度地提高受精成功的机会。精子进化被认为是多配偶物种中精子竞争的主要后果之一,但它也可以通过适应受精地点的环境条件来驱动。强烈的稳定选择限制了可育精子(正常精子)的种内变异,因此多态性有限。在这里,我们分析了利用特征性替代交配行为的雄性之间的生殖形态差异,因此可能存在不同的精子竞争和受精环境条件,在鱿鱼 Loligo bleekeri 中。
大配偶雄性将较小的(平均总长度= 73μm)精子转移到雌性的内部精子储存位置,在输卵管内;而小的 sneaker 雄性将较大的(99μm)精子转移到外部位置,在靠近嘴的精液接收器周围。在游泳速度方面,consort 和 sneaker 精子之间没有观察到显著差异。此外,在精液接收器中,精子优先顺序不受长精子的影响,这表明在 sneaker 雄性中,没有证据表明大精子在争夺精子储存器官中的空间竞争中具有优势。
在这里,我们报告了第一个案例,在鱿鱼 Loligo bleekeri 中,不同大小的雄性产生明显不同的正常精子,这些雄性采用替代交配行为。我们的研究结果没有发现明显的精子二态性是由种内和种间精子竞争驱动的证据。我们提出,替代受精环境的存在,具有不同的特征(即内部或外部),无论是否与精子竞争的影响相结合,都可以驱动精子大小的破坏进化。