Department of Medicine, University of California, San Francisco, California, United States of America.
PLoS Pathog. 2011 Sep;7(9):e1002198. doi: 10.1371/journal.ppat.1002198. Epub 2011 Sep 1.
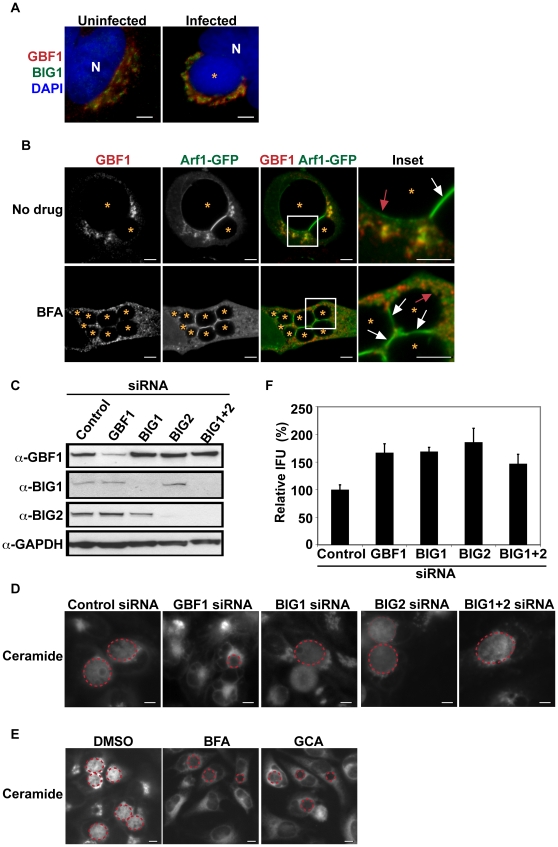
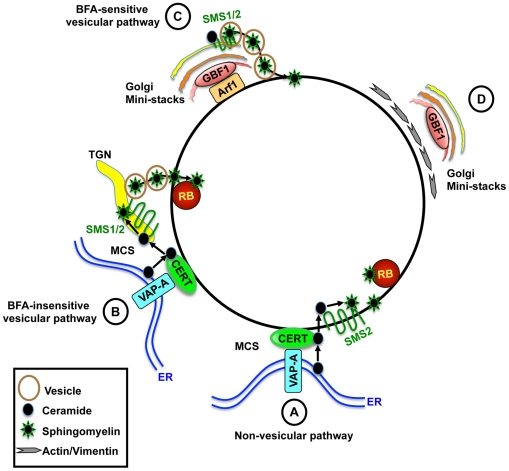
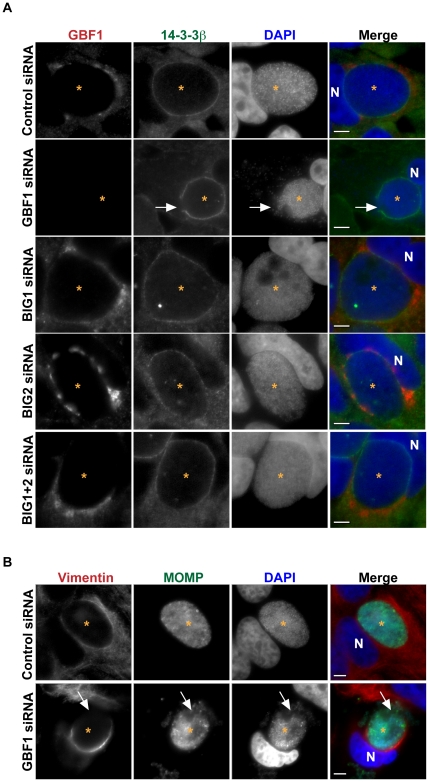
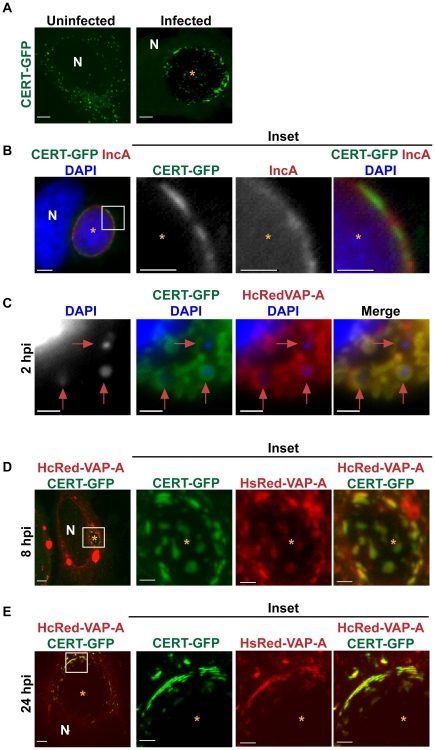
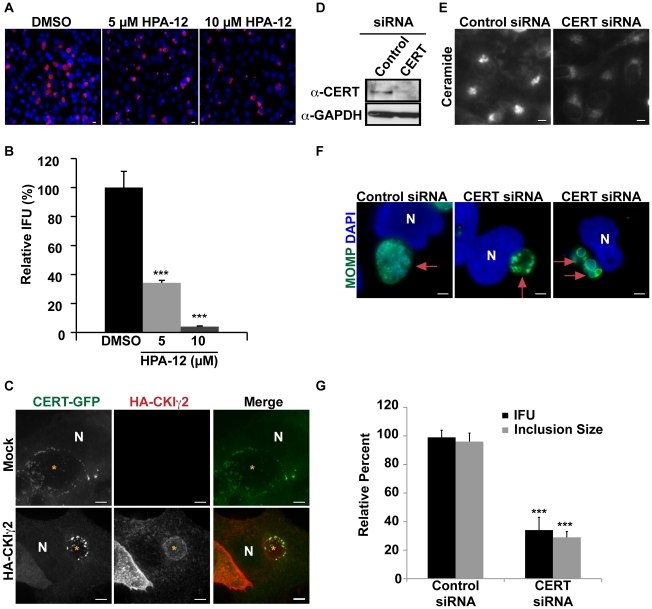
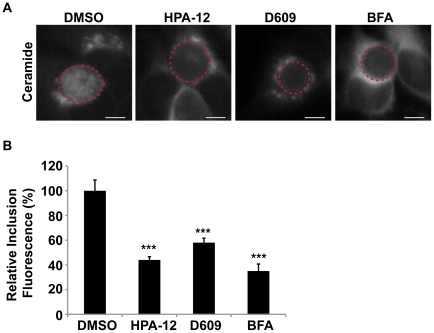
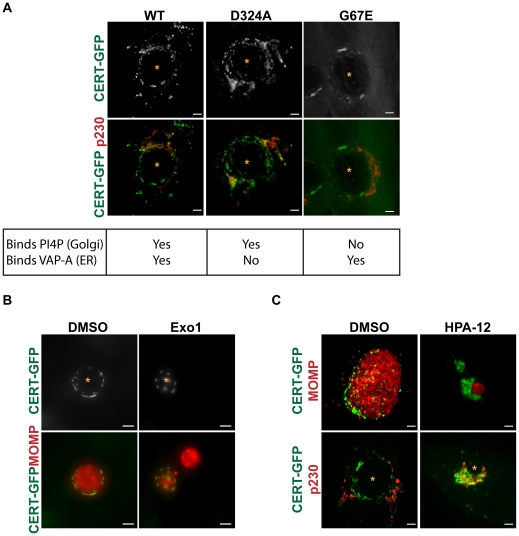
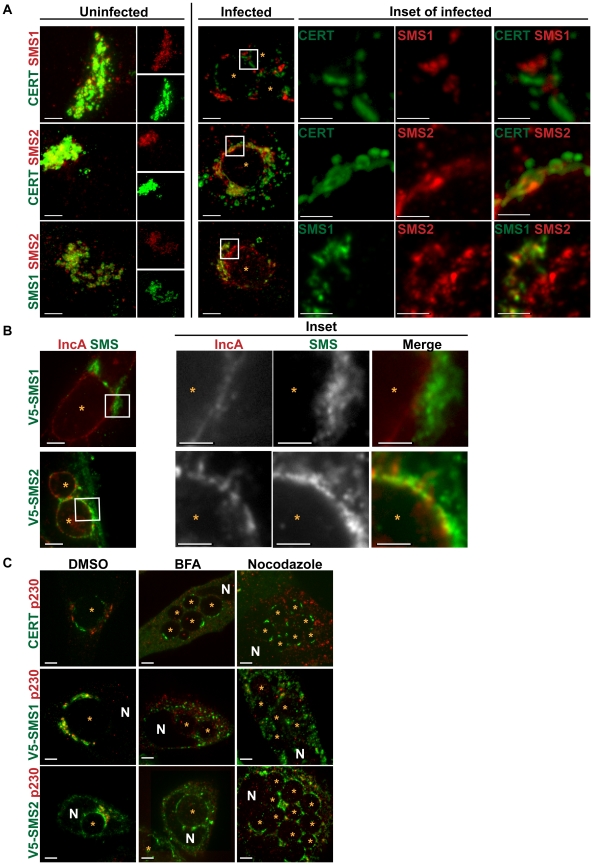
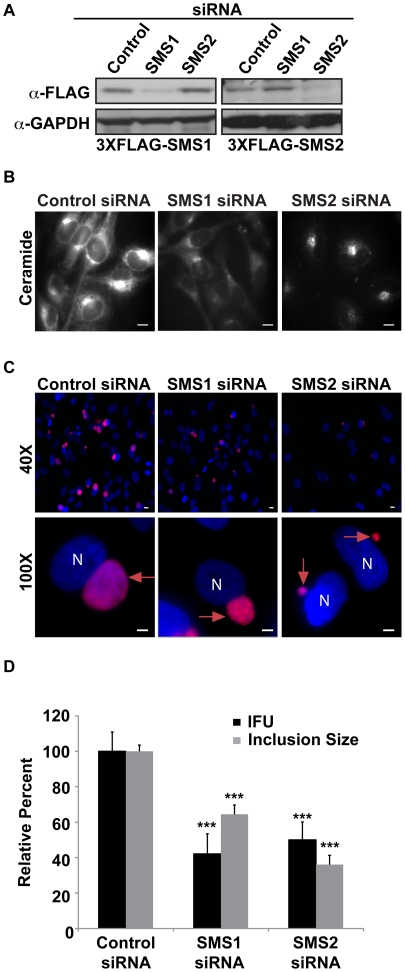
The strain designated Chlamydia trachomatis serovar that was used for experiments in this paper is Chlamydia muridarum, a species closely related to C. trachomatis (and formerly termed the Mouse Pneumonitis strain of C. trachomatis. [corrected]. The obligate intracellular pathogen Chlamydia trachomatis replicates within a membrane-bound inclusion that acquires host sphingomyelin (SM), a process that is essential for replication as well as inclusion biogenesis. Previous studies demonstrate that SM is acquired by a Brefeldin A (BFA)-sensitive vesicular trafficking pathway, although paradoxically, this pathway is dispensable for bacterial replication. This finding suggests that other lipid transport mechanisms are involved in the acquisition of host SM. In this work, we interrogated the role of specific components of BFA-sensitive and BFA-insensitive lipid trafficking pathways to define their contribution in SM acquisition during infection. We found that C. trachomatis hijacks components of both vesicular and non-vesicular lipid trafficking pathways for SM acquisition but that the SM obtained from these separate pathways is being utilized by the pathogen in different ways. We show that C. trachomatis selectively co-opts only one of the three known BFA targets, GBF1, a regulator of Arf1-dependent vesicular trafficking within the early secretory pathway for vesicle-mediated SM acquisition. The Arf1/GBF1-dependent pathway of SM acquisition is essential for inclusion membrane growth and stability but is not required for bacterial replication. In contrast, we show that C. trachomatis co-opts CERT, a lipid transfer protein that is a key component in non-vesicular ER to trans-Golgi trafficking of ceramide (the precursor for SM), for C. trachomatis replication. We demonstrate that C. trachomatis recruits CERT, its ER binding partner, VAP-A, and SM synthases, SMS1 and SMS2, to the inclusion and propose that these proteins establish an on-site SM biosynthetic factory at or near the inclusion. We hypothesize that SM acquired by CERT-dependent transport of ceramide and subsequent conversion to SM is necessary for C. trachomatis replication whereas SM acquired by the GBF1-dependent pathway is essential for inclusion growth and stability. Our results reveal a novel mechanism by which an intracellular pathogen redirects SM biosynthesis to its replicative niche.
本文实验中使用的衣原体血清型菌株命名为沙眼衣原体鼠亚种,它是与沙眼衣原体密切相关的一种物种(以前被称为沙眼衣原体鼠肺炎株。[纠正]。专性细胞内病原体沙眼衣原体在膜结合包涵体内复制,该包涵体获得宿主神经鞘磷脂(SM),这一过程对于复制和包涵体发生都是必不可少的。先前的研究表明,SM 通过布雷菲德菌素 A(BFA)敏感的小泡运输途径获得,尽管矛盾的是,该途径对于细菌复制是可有可无的。这一发现表明,其他脂质运输机制参与了宿主 SM 的获取。在这项工作中,我们研究了 BFA 敏感和 BFA 不敏感的脂质运输途径中的特定成分的作用,以确定它们在感染过程中对 SM 摄取的贡献。我们发现,沙眼衣原体劫持了小泡和非小泡脂质运输途径的成分来获取 SM,但病原体以不同的方式利用从这些不同途径获得的 SM。我们表明,沙眼衣原体选择性地仅劫持三种已知的 BFA 靶标之一,GBF1,即早期分泌途径中 Arf1 依赖性小泡运输的调节剂,用于小泡介导的 SM 摄取。SM 摄取的 Arf1/GBF1 依赖性途径对于包涵体膜的生长和稳定性是必不可少的,但对于细菌复制是不必要的。相反,我们表明,沙眼衣原体劫持 CERT,一种脂质转移蛋白,它是神经酰胺(SM 的前体)非小泡内质网到反式高尔基体运输的关键组成部分,用于沙眼衣原体复制。我们证明沙眼衣原体招募 CERT、其内质网结合伴侣 VAP-A 和 SM 合酶 SMS1 和 SMS2 到包涵体,并提出这些蛋白在包涵体或其附近建立一个现场 SM 生物合成工厂。我们假设,由 CERT 依赖性的神经酰胺运输和随后转化为 SM 所获得的 SM 对于沙眼衣原体的复制是必要的,而由 GBF1 依赖性途径获得的 SM 对于包涵体的生长和稳定性是必要的。我们的结果揭示了一种新的机制,即细胞内病原体将 SM 生物合成重新定向到其复制位。