Department of Molecular Biophysics, Faculty of Biochemistry, Biophysics and Biotechnology, Jagiellonian University, Kraków, Poland.
Biochemistry. 2012 Jan 31;51(4):829-35. doi: 10.1021/bi2016316. Epub 2012 Jan 17.
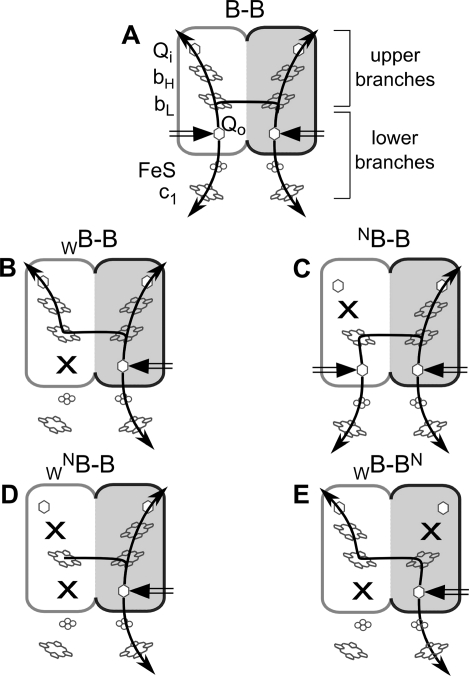
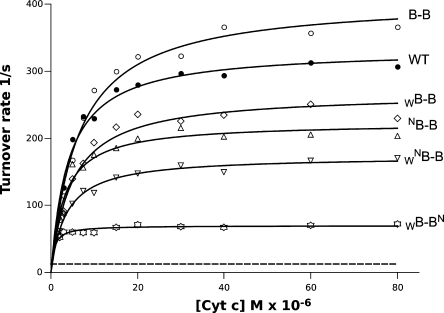
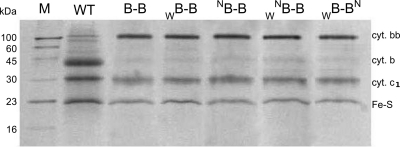
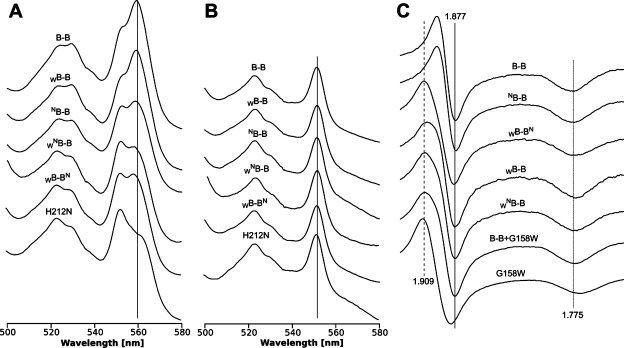
Homodimeric structure of cytochrome bc₁, a common component of biological energy conversion systems, builds in four catalytic quinone oxidation/reduction sites and four chains of cofactors (branches) that, connected by a centrally located bridge, form a symmetric H-shaped electron transfer system. The mechanism of operation of this complex system is under constant debate. Here, we report on isolation and enzymatic examination of cytochrome bc₁-like complexes containing fused cytochrome b subunits in which asymmetrically introduced mutations inactivated individual branches in various combinations. The structural asymmetry of those forms was confirmed spectroscopically. All the asymmetric forms corresponding to cytochrome bc₁ with partial or full inactivation of one monomer retain high enzymatic activity but at the same time show a decrease in the maximum turnover rate by a factor close to 2. This strongly supports the model assuming independent operation of monomers. The cross-inactivated form corresponding to cytochrome bc₁ with disabled complementary parts of each monomer retains the enzymatic activity at the level that, for the first time on isolated from membranes and purified to homogeneity preparations, demonstrates that intermonomer electron transfer through the bridge effectively sustains the enzymatic turnover. The results fully support the concept that electrons freely distribute between the four catalytic sites of a dimer and that any path connecting the catalytic sites on the opposite sides of the membrane is enzymatically competent. The possibility to examine enzymatic properties of isolated forms of asymmetric complexes constructed using the cytochrome b fusion system extends the array of tools available for investigating the engineering of dimeric cytochrome bc₁ from the mechanistic and physiological perspectives.
细胞色素 bc₁的同源二聚体结构是生物能量转换系统的常见组成部分,它构建了四个催化醌氧化/还原位点和四个辅因子链(分支),这些分支通过中央桥连接,形成对称的 H 形电子传递系统。这个复杂系统的工作机制一直存在争议。在这里,我们报告了含有融合细胞色素 b 亚基的细胞色素 bc₁样复合物的分离和酶学研究,其中在不对称引入的突变中,以各种组合失活了单个分支。这些形式的结构不对称性通过光谱学得到了证实。所有对应于细胞色素 bc₁的不对称形式,其单体的一部分或全部失活,保留了高酶活性,但同时最大周转率降低了接近 2 的因子。这强烈支持了单体独立运作的模型。对应于每个单体互补部分失活的细胞色素 bc₁的交叉失活形式保留了酶活性,这在从膜中分离并纯化为均质制剂的首次实验中证明,桥接处的单体间电子转移有效地维持了酶的周转率。结果完全支持了这样的概念,即电子在二聚体的四个催化位点之间自由分配,并且任何连接膜两侧催化位点的路径在酶学上都是有效的。使用细胞色素 b 融合系统构建不对称复合物的分离形式的酶学性质的研究扩展了用于从机制和生理角度研究二聚体细胞色素 bc₁工程的工具组合。