Department of Biology, York College of the City University of New York, Jamaica, New York 11451, USA.
Mob DNA. 2012 Jan 26;3(1):1. doi: 10.1186/1759-8753-3-1.
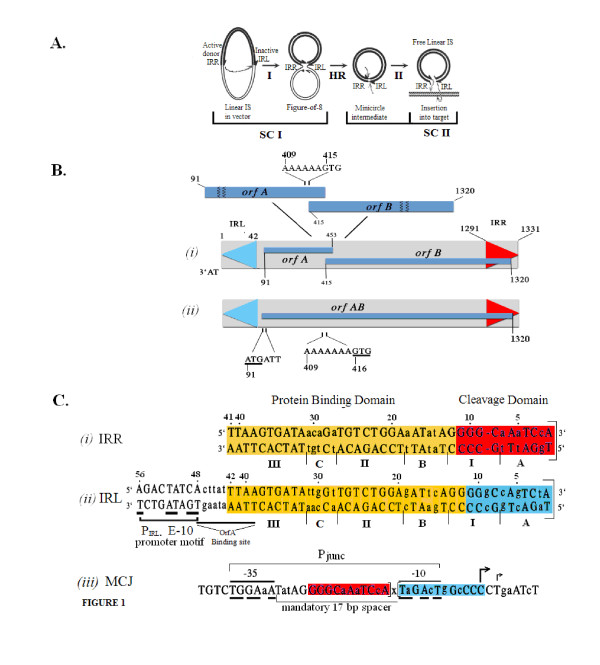
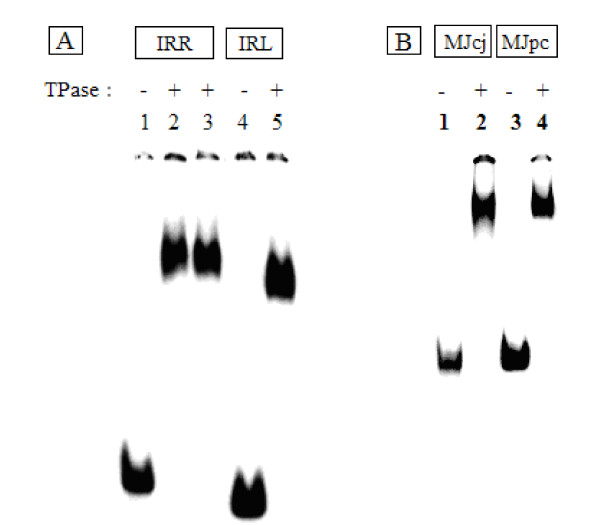
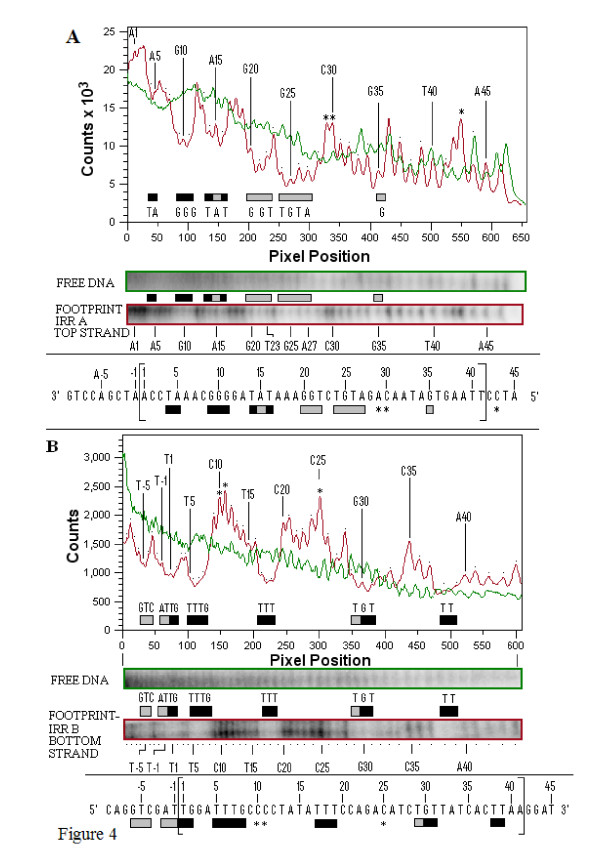
Transposition in IS3, IS30, IS21 and IS256 insertion sequence (IS) families utilizes an unconventional two-step pathway. A figure-of-eight intermediate in Step I, from asymmetric single-strand cleavage and joining reactions, is converted into a double-stranded minicircle whose junction (the abutted left and right ends) is the substrate for symmetrical transesterification attacks on target DNA in Step II, suggesting intrinsically different synaptic complexes (SC) for each step. Transposases of these ISs bind poorly to cognate DNA and comparative biophysical analyses of SC I and SC II have proven elusive. We have prepared a native, soluble, active, GFP-tagged fusion derivative of the IS2 transposase that creates fully formed complexes with single-end and minicircle junction (MCJ) substrates and used these successfully in hydroxyl radical footprinting experiments.
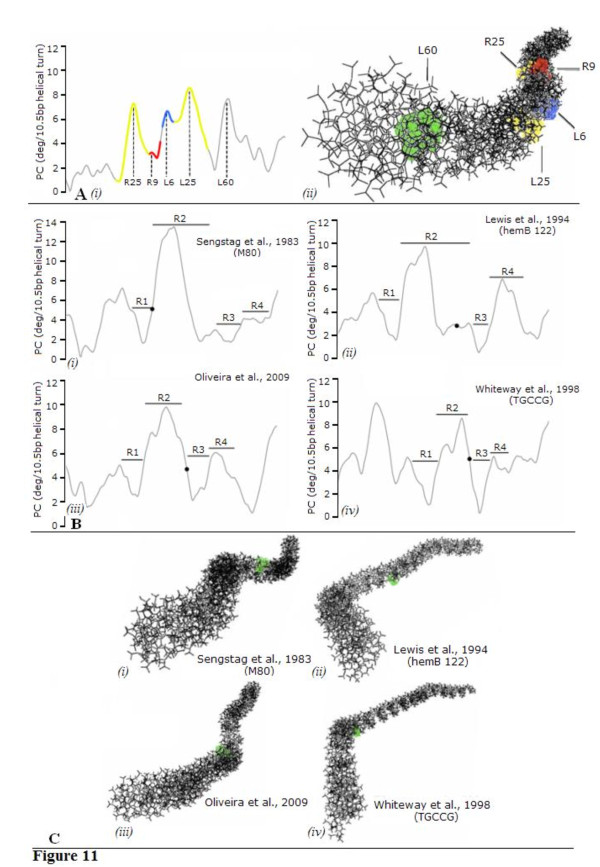
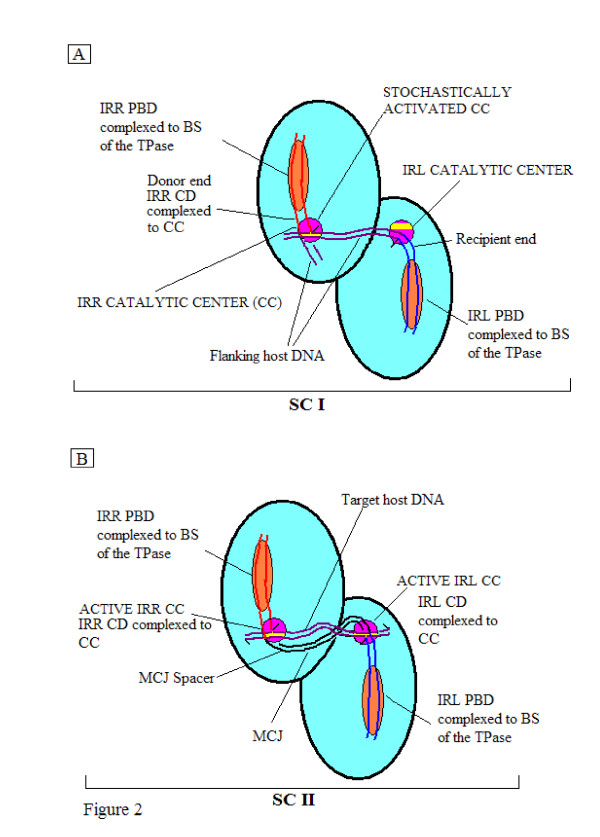
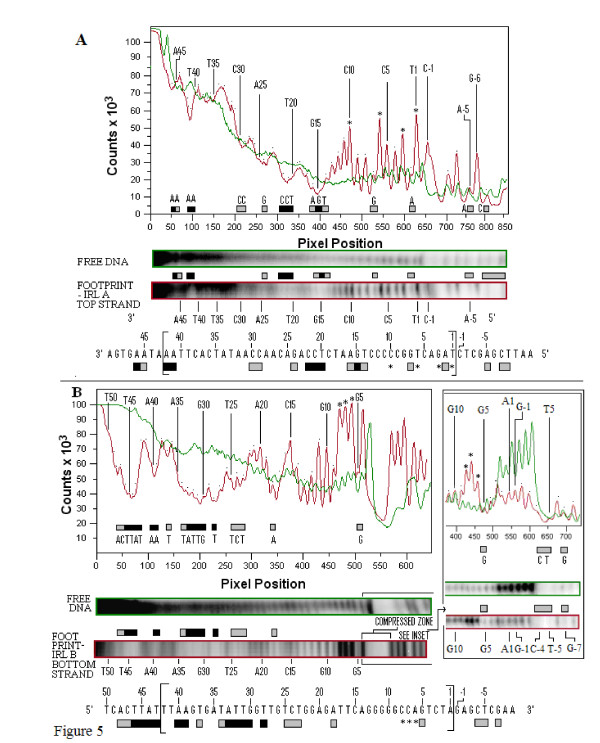
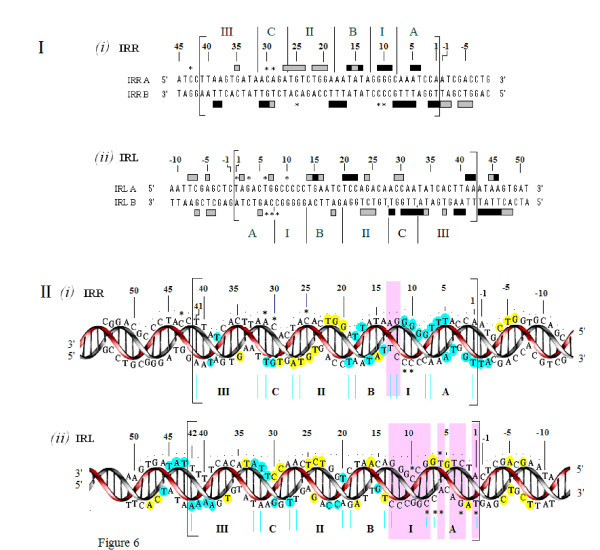
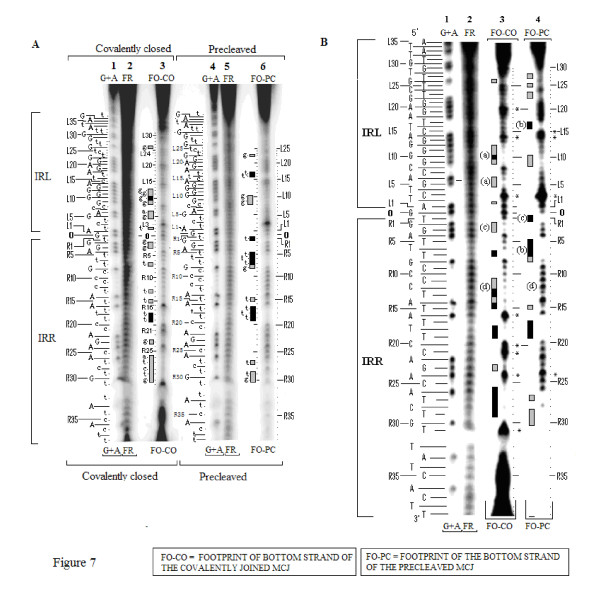
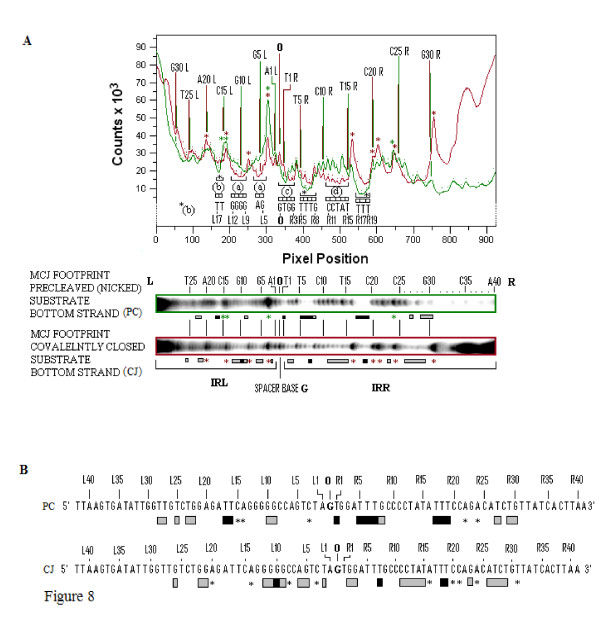
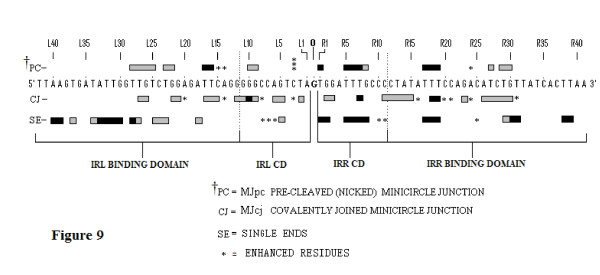
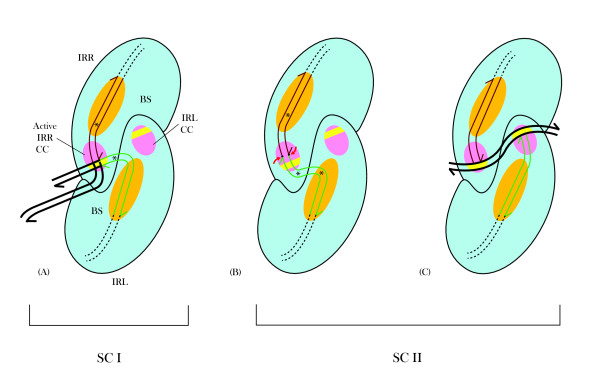
In IS2, Step I reactions are physically and chemically asymmetric; the left imperfect, inverted repeat (IRL), the exclusive recipient end, lacks donor function. In SC I, different protection patterns of the cleavage domains (CDs) of the right imperfect inverted repeat (IRR; extensive in cis) and IRL (selective in trans) at the single active cognate IRR catalytic center (CC) are related to their donor and recipient functions. In SC II, extensive binding of the IRL CD in trans and of the abutted IRR CD in cis at this CC represents the first phase of the complex. An MCJ substrate precleaved at the 3' end of IRR revealed a temporary transition state with the IRL CD disengaged from the protein. We propose that in SC II, sequential 3' cleavages at the bound abutted CDs trigger a conformational change, allowing the IRL CD to complex to its cognate CC, producing the second phase. Corroborating data from enhanced residues and curvature propensity plots suggest that CD to CD interactions in SC I and SC II require IRL to assume a bent structure, to facilitate binding in trans.
Different transpososomes are assembled in each step of the IS2 transposition pathway. Recipient versus donor end functions of the IRL CD in SC I and SC II and the conformational change in SC II that produces the phase needed for symmetrical IRL and IRR donor attacks on target DNA highlight the differences.
转座在 IS3、IS30、IS21 和 IS256 插入序列(IS)家族中利用非常规的两步途径。第一步中的“8”字形中间体来自不对称单链切割和连接反应,转化为双链的迷你环,其连接点(左右对齐的末端)是第二步中目标 DNA 上对称转酯攻击的底物,表明每个步骤都有内在不同的突触复合物(SC)。这些 IS 的转座酶与同源 DNA 的结合能力较差,并且 SC I 和 SC II 的比较生物物理分析一直难以捉摸。我们已经制备了 IS2 转座酶的天然、可溶性、活性 GFP 标记融合衍生物,该衍生物可与单端和迷你环连接(MCJ)底物形成完全形成的复合物,并在羟基自由基足迹实验中成功使用这些复合物。
在 IS2 中,第一步反应在物理和化学上是不对称的;左边不完美的反向重复(IRL),唯一的接受末端,缺乏供体功能。在 SC I 中,在单活性同源 IRR 催化中心(CC)处,右不完美反向重复(IRR;顺式广泛)和 IRL(反式选择性)的切割结构域(CD)的不同保护模式与它们的供体和受体功能有关。在 SC II 中,IRL CD 在反式中的广泛结合和 abutted IRR CD 在顺式中的结合在该 CC 处代表了复合物的第一阶段。在 IRR 的 3'末端预先切割的 MCJ 底物揭示了一个暂时的过渡态,其中 IRL CD 与蛋白质脱离。我们提出,在 SC II 中,结合的 abutted CDs 的连续 3'切割触发构象变化,允许 IRL CD 与其同源 CC 复合,产生第二阶段。来自增强残基和曲率倾向图的佐证数据表明,SC I 和 SC II 中的 CD 到 CD 相互作用需要 IRL 采用弯曲结构,以促进反式结合。
IS2 转位途径的每一步都组装了不同的转座体。在 SC I 和 SC II 中,IRL CD 的受体与供体功能以及在 SC II 中产生用于对称 IRL 和 IRR 供体攻击目标 DNA 的所需相位的构象变化突出了这些差异。