Department of Physics, University of Genova, Genova, Italy.
PLoS One. 2013 Jun 12;8(6):e66144. doi: 10.1371/journal.pone.0066144. Print 2013.
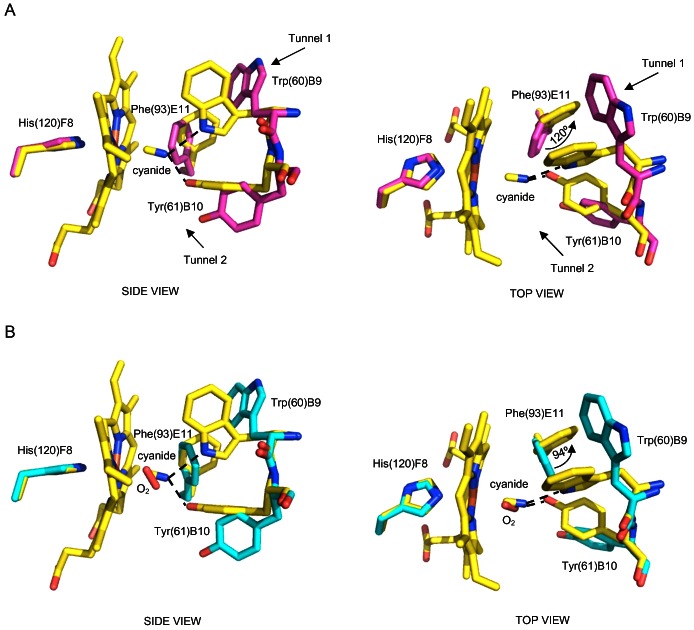
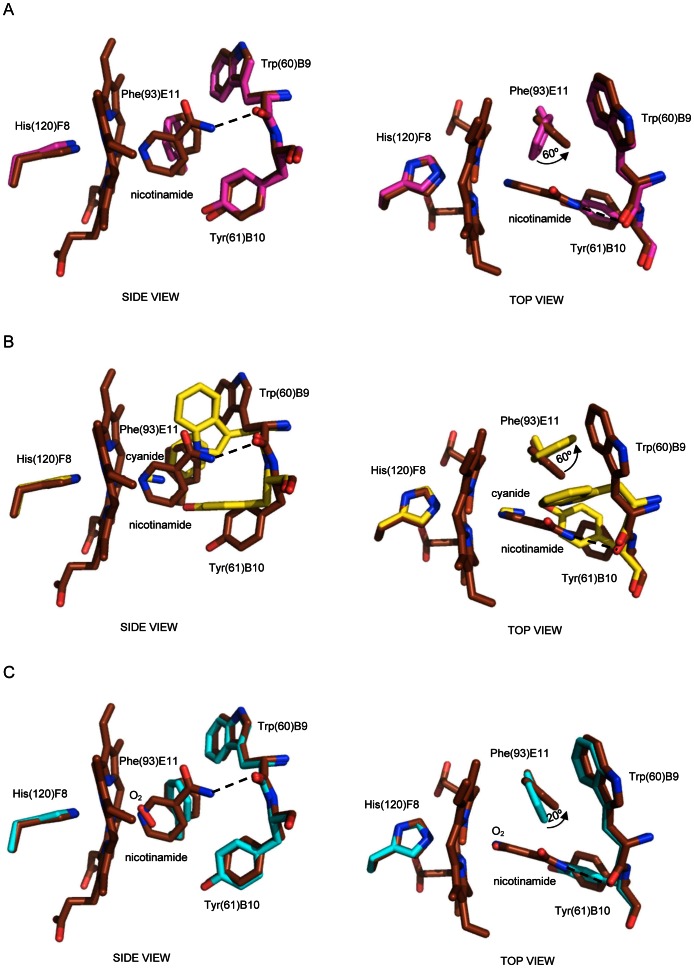
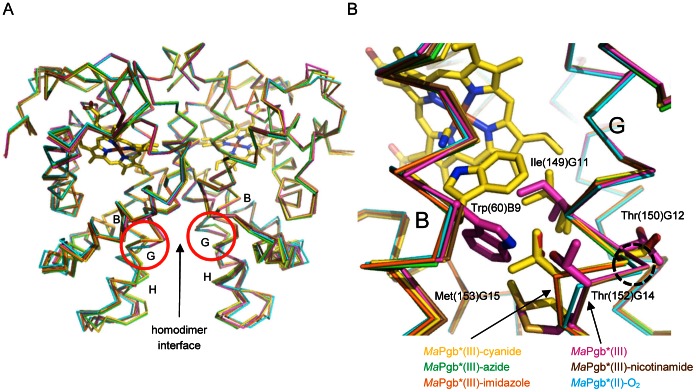
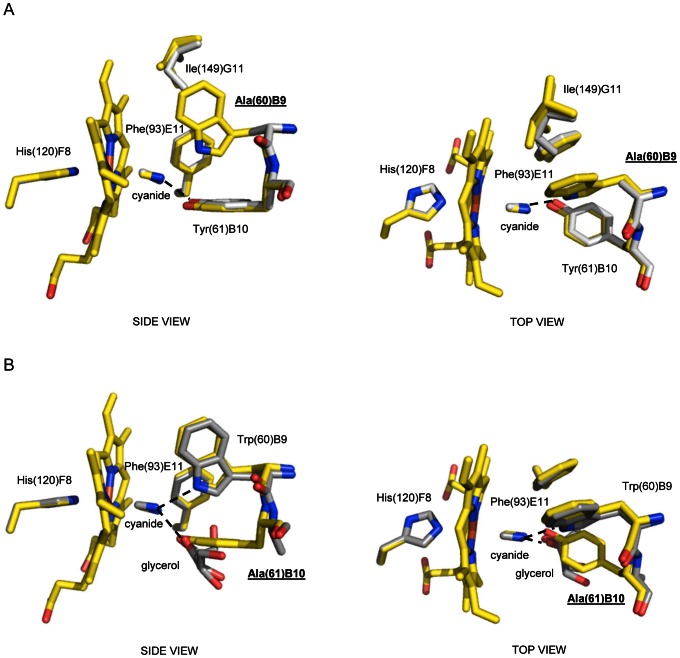
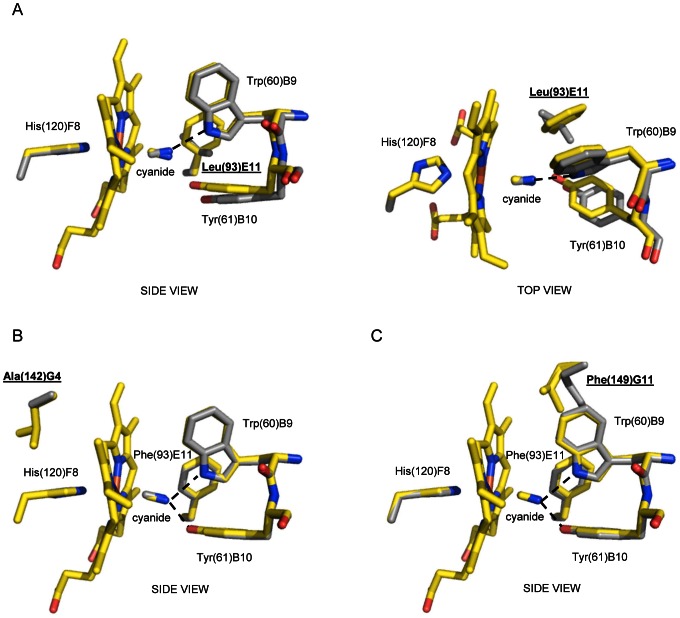
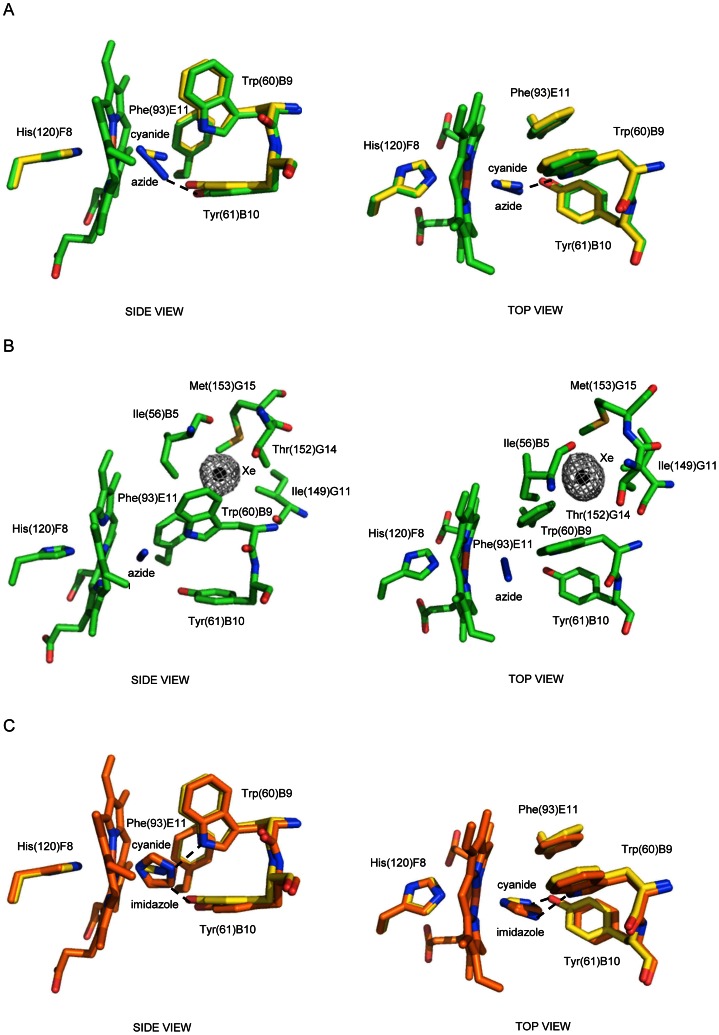
Protoglobin from Methanosarcina acetivorans C2A (MaPgb), a strictly anaerobic methanogenic Archaea, is a dimeric haem-protein whose biological role is still unknown. As other globins, protoglobin can bind O2, CO and NO reversibly in vitro, but it displays specific functional and structural properties within members of the hemoglobin superfamily. CO binding to and dissociation from the haem occurs through biphasic kinetics, which arise from binding to (and dissociation from) two distinct tertiary states in a ligation-dependent equilibrium. From the structural viewpoint, protoglobin-specific loops and a N-terminal extension of 20 residues completely bury the haem within the protein matrix. Thus, access of small ligand molecules to the haem is granted by two apolar tunnels, not common to other globins, which reach the haem distal site from locations at the B/G and B/E helix interfaces. Here, the roles played by residues Trp(60)B9, Tyr(61)B10 and Phe(93)E11 in ligand recognition and stabilization are analyzed, through crystallographic investigations on the ferric protein and on selected mutants. Specifically, protein structures are reported for protoglobin complexes with cyanide, with azide (also in the presence of Xenon), and with more bulky ligands, such as imidazole and nicotinamide. Values of the rate constant for cyanide dissociation from ferric MaPgb-cyanide complexes have been correlated to hydrogen bonds provided by Trp(60)B9 and Tyr(61)B10 that stabilize the haem-Fe(III)-bound cyanide. We show that protoglobin can strikingly reshape, in a ligand-dependent way, the haem distal site, where Phe(93)E11 acts as ligand sensor and controls accessibility to the haem through the tunnel system by modifying the conformation of Trp(60)B9.
产甲烷八叠球菌 C2A(MaPgb)的原球蛋白是一种严格厌氧的产甲烷古菌中的二聚血红素蛋白,其生物学功能尚不清楚。与其他球蛋白一样,原球蛋白可以在体外可逆地结合 O2、CO 和 NO,但它在血红蛋白超家族成员中表现出特定的功能和结构特性。CO 与血红素的结合和解离呈两相动力学,这是由于在配体依赖性平衡中与两个不同的三级状态结合(和解离)而产生的。从结构角度来看,原球蛋白特有的环和 20 个残基的 N 端延伸将血红素完全埋藏在蛋白质基质中。因此,小分子配体分子进入血红素的通道是通过两个非极性隧道实现的,这些隧道不同于其他球蛋白,它们从 B/G 和 B/E 螺旋界面的位置到达血红素的远端部位。在这里,通过对铁蛋白和选定突变体的晶体学研究,分析了残基 Trp(60)B9、Tyr(61)B10 和 Phe(93)E11 在配体识别和稳定中的作用。具体来说,报告了原球蛋白与氰化物、叠氮化物(也存在氙气)以及更庞大配体(如咪唑和烟酰胺)复合物的蛋白质结构。铁 MaPgb-氰化物复合物中氰化物从铁(III)解离的速率常数与 Trp(60)B9 和 Tyr(61)B10 提供的氢键相关,这些氢键稳定了血红素-Fe(III)-结合的氰化物。我们表明,原球蛋白可以通过改变 Trp(60)B9 的构象,以配体依赖的方式显著重塑血红素的远端部位,其中 Phe(93)E11 作为配体传感器,并通过隧道系统控制血红素的可及性。