Department of Biological Sciences, Louisiana State University , Baton Rouge, Louisiana 70803, United States.
Biochemistry. 2014 Jul 15;53(27):4368-80. doi: 10.1021/bi500219t. Epub 2014 Jul 1.
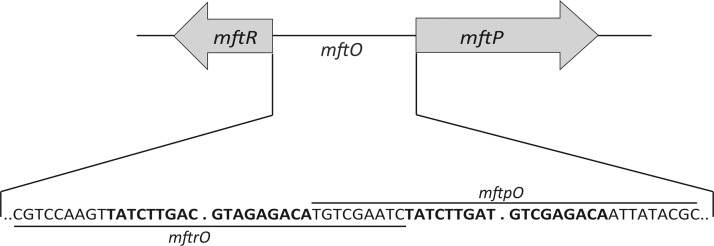
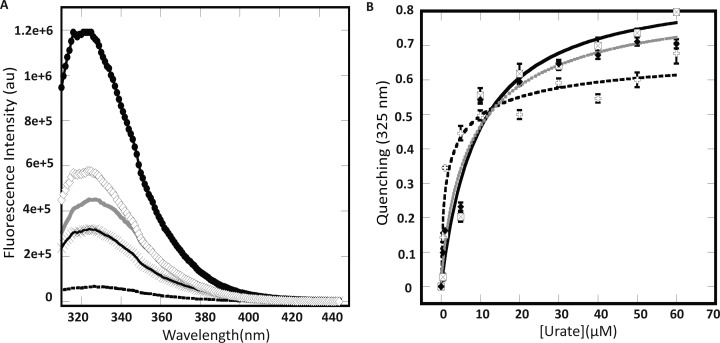
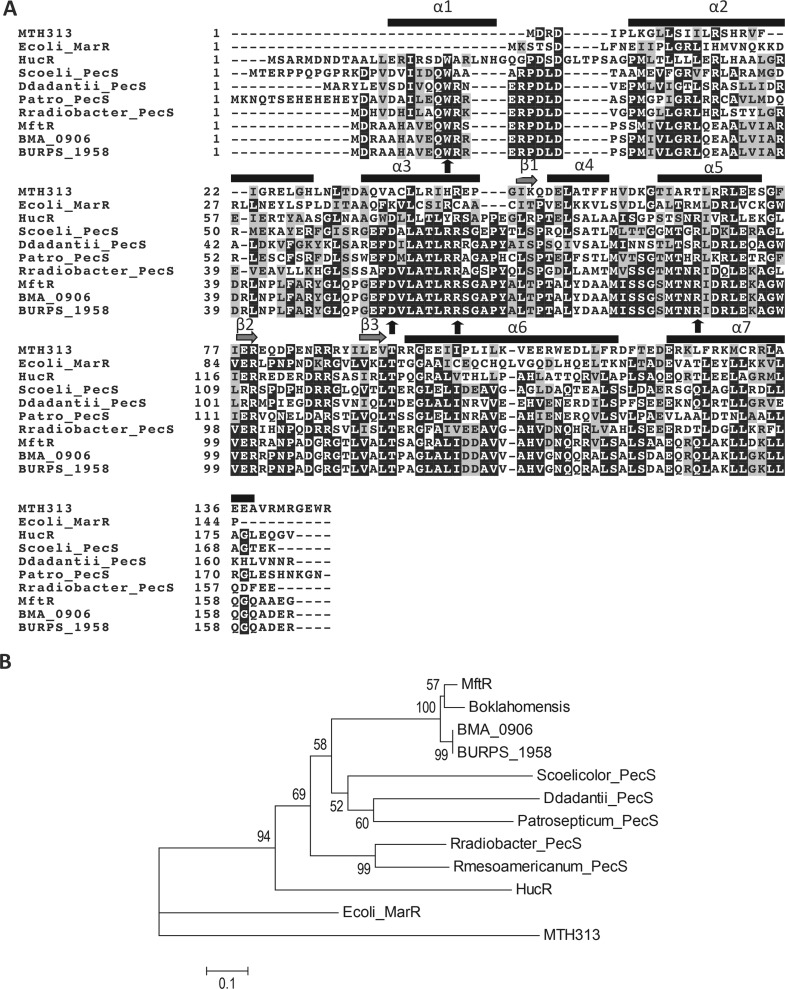
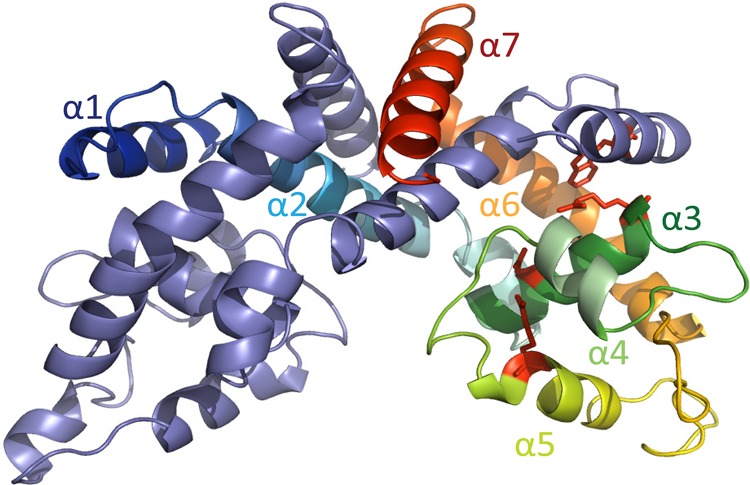
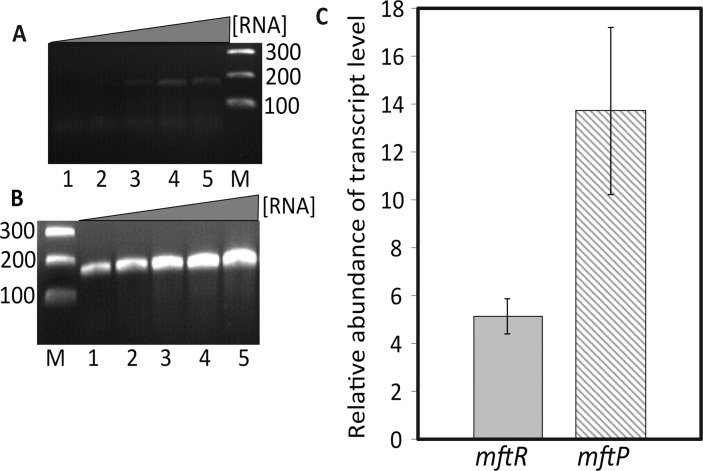
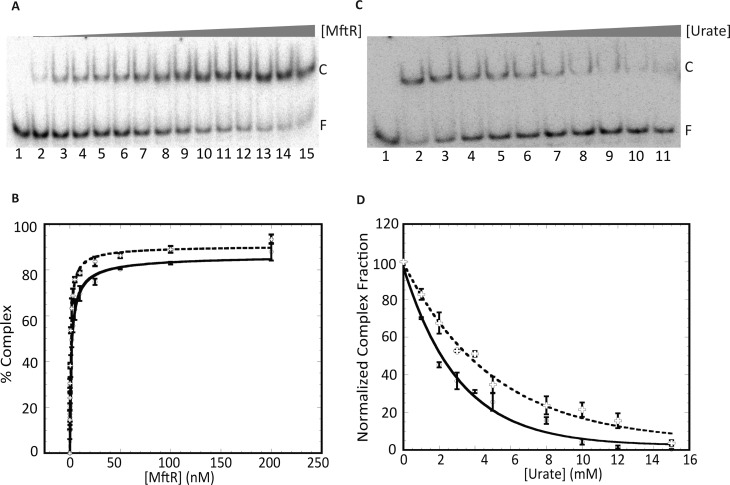
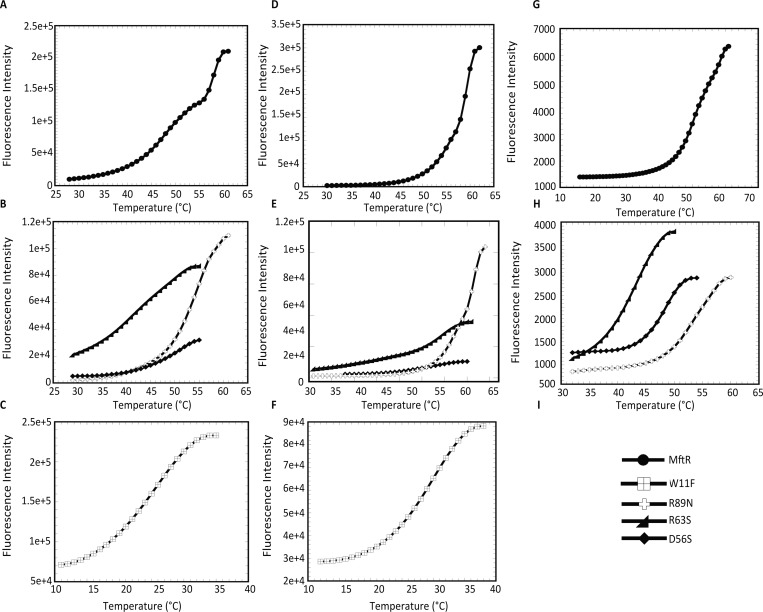
Members of the multiple antibiotic resistance regulator (MarR) family often regulate gene activity by responding to a specific ligand. In the absence of ligand, most MarR proteins function as repressors, while ligand binding causes attenuated DNA binding and therefore increased gene expression. Previously, we have shown that urate is a ligand for MftR (major facilitator transport regulator), which is encoded by the soil bacterium Burkholderia thailandensis. We show here that both mftR and the divergently oriented gene mftP encoding a major facilitator transport protein are upregulated in the presence of urate. MftR binds two cognate sites in the mftR-mftP intergenic region with equivalent affinity and sensitivity to urate. Mutagenesis of four conserved residues previously reported to be involved in urate binding to Deinococcus radiodurans HucR and Rhizobium radiobacter PecS significantly reduced protein stability and DNA binding affinity but not ligand binding. These data suggest that residues equivalent to those implicated in ligand binding to HucR and PecS serve structural roles and that MftR relies on distinct residues for ligand binding. MftR exhibits a two-step melting transition suggesting independent unfolding of the dimerization and DNA-binding regions; urate binding or mutations in the predicted ligand-binding sites result in one-step unfolding transitions. We suggest that MftR binds the ligand in a cleft between the DNA-binding lobes and the dimer interface but that the mechanism of ligand-mediated attenuation of DNA binding differs from that proposed for other urate-responsive MarR homologues. Since DNA binding by MftR is attenuated at 37 °C, our data also suggest that MftR responds to both ligand and a thermal upshift by attenuated DNA binding and upregulation of the genes under its control.
多抗生素耐药调节蛋白(MarR)家族的成员通常通过响应特定配体来调节基因活性。在没有配体的情况下,大多数 MarR 蛋白作为抑制剂发挥作用,而配体结合会导致 DNA 结合减弱,从而增加基因表达。先前,我们已经表明尿酸是 MftR(主要易化转运调节剂)的配体,MftR 由土壤细菌伯克霍尔德氏菌编码。我们在这里表明,在尿酸存在的情况下,mftR 和编码主要易化转运蛋白的定向基因 mftP 都被上调。MftR 以相等的亲和力和对尿酸的敏感性结合在 mftR-mftP 基因间区的两个同源位点上。先前报道的涉及与 Deinococcus radiodurans HucR 和 Rhizobium radiobacter PecS 结合的尿酸的四个保守残基的突变显著降低了蛋白质稳定性和 DNA 结合亲和力,但不降低配体结合。这些数据表明,与涉及与 HucR 和 PecS 结合的配体的残基等效的残基起结构作用,并且 MftR 依赖于不同的残基进行配体结合。MftR 表现出两步熔融转变,表明二聚化和 DNA 结合区域的独立展开;尿酸结合或预测配体结合位点的突变导致一步展开转变。我们认为 MftR 在 DNA 结合结构域和二聚体界面之间的裂隙中结合配体,但配体介导的 DNA 结合减弱的机制与其他尿酸响应性 MarR 同源物提出的机制不同。由于 MftR 在 37°C 时 DNA 结合减弱,我们的数据还表明 MftR 通过减弱 DNA 结合和受其控制的基因的上调来响应配体和热移位。