Onel Melis, Sumbul Fidan, Liu Jin, Nussinov Ruth, Haliloglu Turkan
Polymer Research Center and Chemical Engineering Department, Bogazici University, Istanbul, Turkey.
Department of Pharmaceutical Sciences, University of North Texas System College of Pharmacy, University of North Texas Health Science Center, Fort Worth, TX, U.S.A.
Biochem J. 2017 Feb 20;474(5):781-795. doi: 10.1042/BCJ20160748.
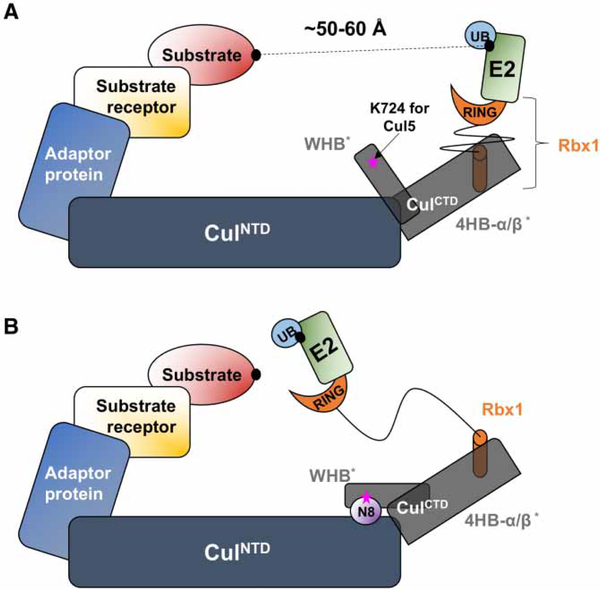
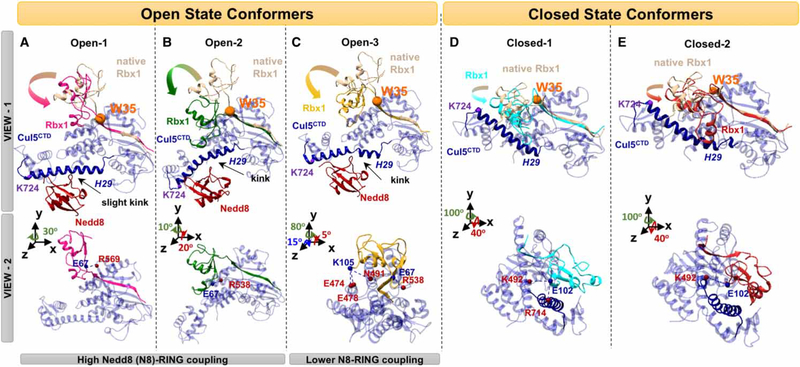
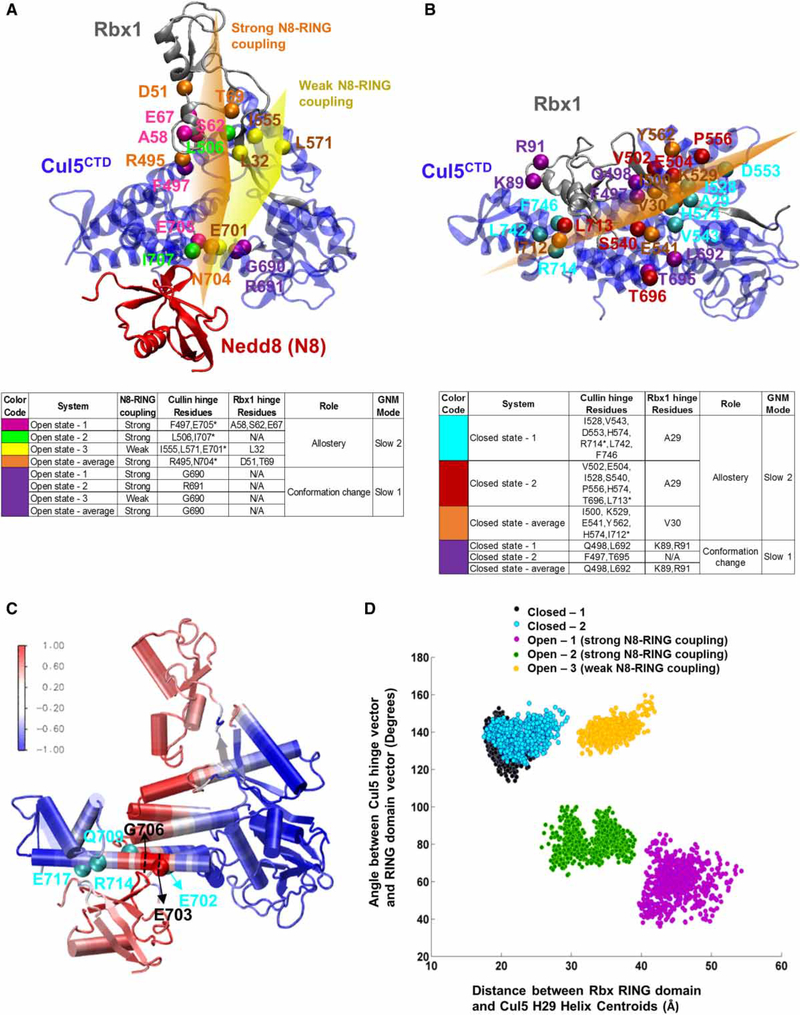
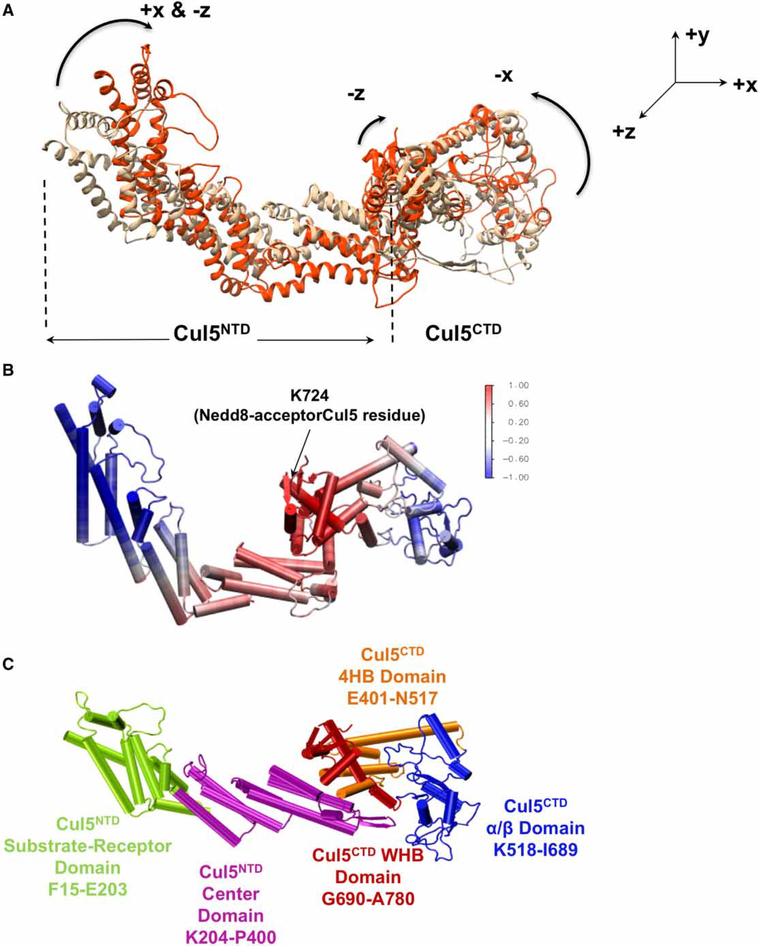
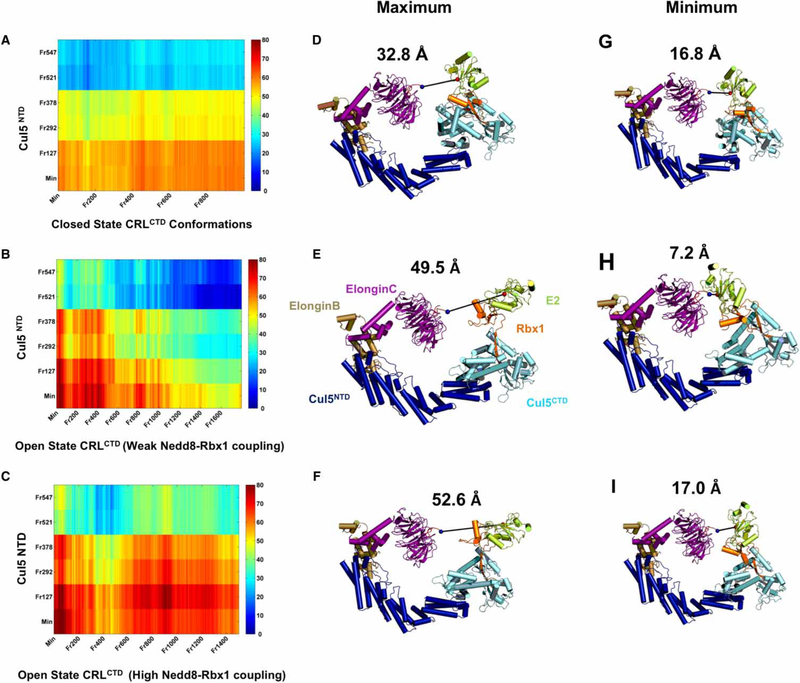
Conjugation of Nedd8 (neddylation) to Cullins (Cul) in Cul-RING E3 ligases (CRLs) stimulates ubiquitination and polyubiquitination of protein substrates. CRL is made up of two Cul-flanked arms: one consists of the substrate-binding and adaptor proteins and the other consists of E2 and Ring-box protein (Rbx). Polyubiquitin chain length and topology determine the substrate fate. Here, we ask how polyubiquitin chains are accommodated in the limited space available between the two arms and what determines the polyubiquitin linkage topology. We focus on Cul5 and Rbx1 in three states: before Cul5 neddylation (closed state), after neddylation (open state), and after deneddylation, exploiting molecular dynamics simulations and the Gaussian Network Model. We observe that regulation of substrate ubiquitination and polyubiquitination takes place through Rbx1 rotations, which are controlled by Nedd8-Rbx1 allosteric communication. Allosteric propagation proceeds from Nedd8 via Cul5 dynamic hinges and hydrogen bonds between the C-terminal domain of Cul5 (Cul5) and Rbx1 (Cul5 residues R538/R569 and Rbx1 residue E67, or Cul5 E474/E478/N491 and Rbx1 K105). Importantly, at each ubiquitination step (homogeneous or heterogeneous, linear or branched), the polyubiquitin linkages fit into the distances between the two arms, and these match the inherent CRL conformational tendencies. Hinge sites may constitute drug targets.
在Cul-RING E3连接酶(CRL)中,Nedd8与Cullins(Cul)的缀合(Neddylation)刺激蛋白质底物的泛素化和多聚泛素化。CRL由两个以Cul为侧翼的臂组成:一个由底物结合蛋白和衔接蛋白组成,另一个由E2和环框蛋白(Rbx)组成。多聚泛素链的长度和拓扑结构决定底物的命运。在这里,我们研究多聚泛素链如何在两个臂之间有限的空间中容纳,以及是什么决定了多聚泛素连接的拓扑结构。我们利用分子动力学模拟和高斯网络模型,聚焦于处于三种状态的Cul5和Rbx1:Cul5进行Neddylation修饰之前(闭合状态)、修饰之后(开放状态)以及去Neddylation修饰之后。我们观察到底物泛素化和多聚泛素化的调节是通过Rbx1的旋转进行的,而Rbx1的旋转由Nedd8-Rbx1变构通讯控制。变构传播从Nedd8经由Cul5动态铰链以及Cul5(Cul5)的C末端结构域与Rbx1(Cul5的R538/R569残基和Rbx1的E67残基,或Cul5的E474/E478/N491残基和Rbx1的K105残基)之间的氢键进行。重要的是,在每个泛素化步骤(均一或不均一、线性或分支)中,多聚泛素连接适合于两个臂之间的距离,并且这些距离与CRL固有的构象倾向相匹配。铰链位点可能构成药物靶点。