Institute for Zoology/Developmental Biology, Biocenter, University of Cologne, Köln, Germany.
Department of Zoology, University of Cambridge, Cambridge, United Kingdom.
Elife. 2019 Oct 1;8:e47346. doi: 10.7554/eLife.47346.
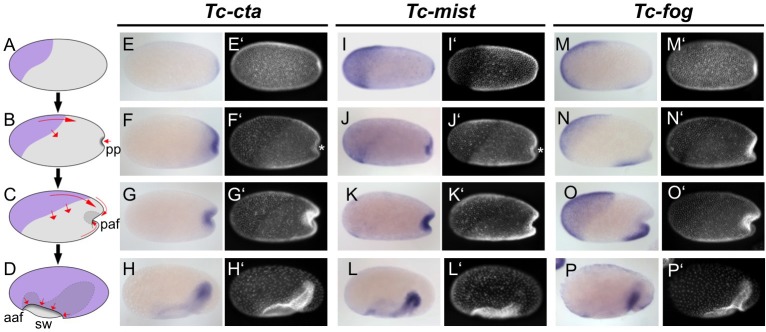
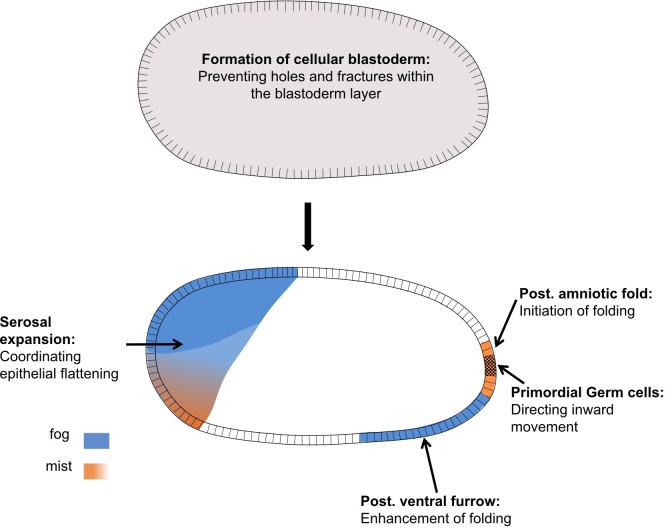
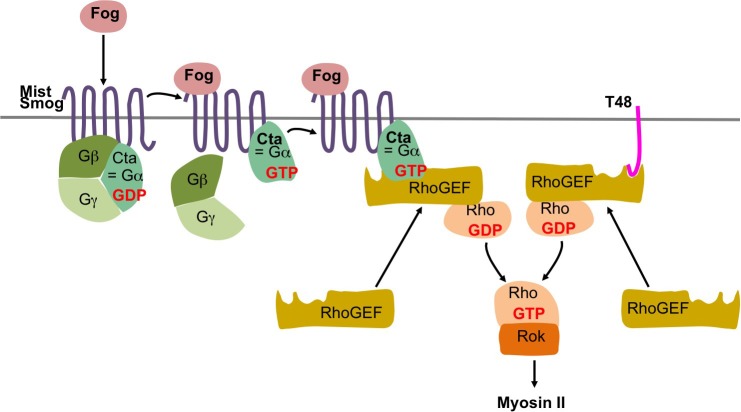
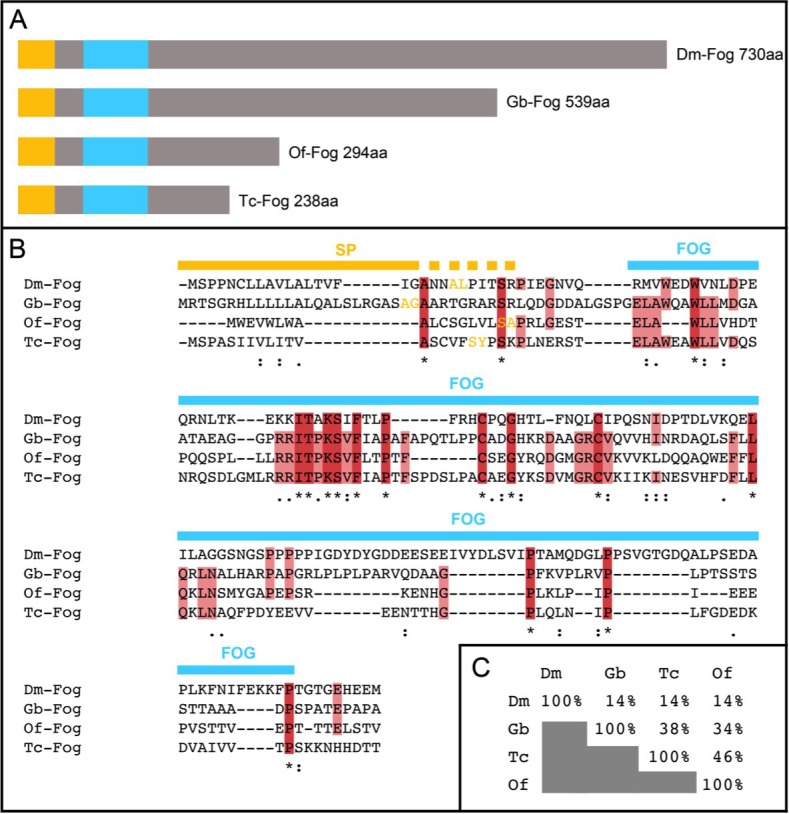
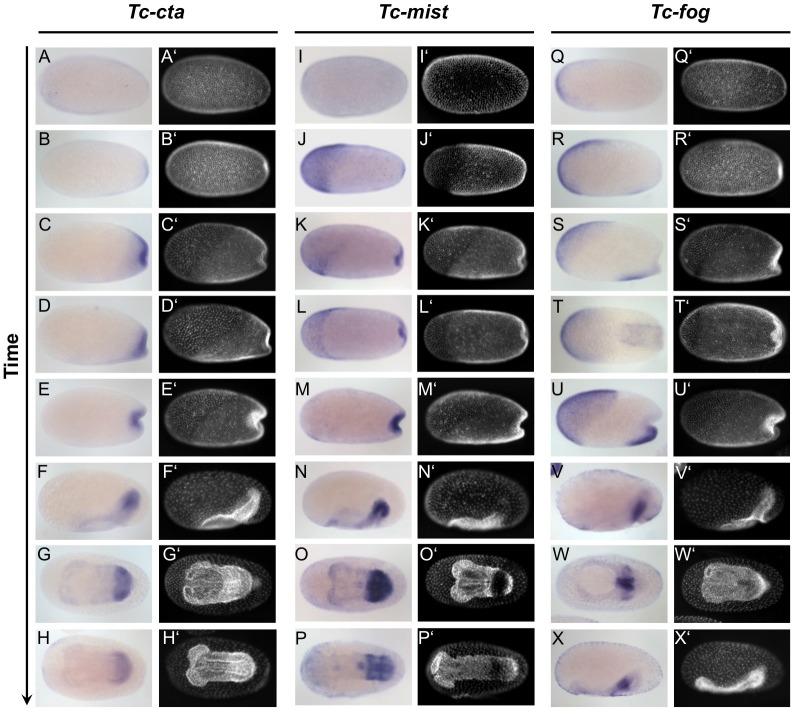
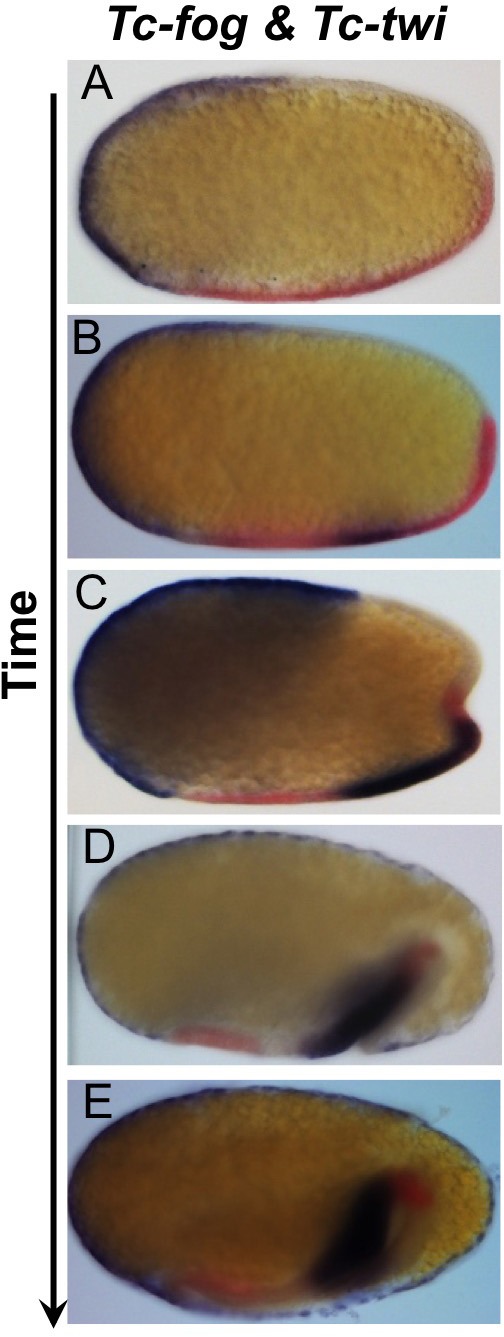
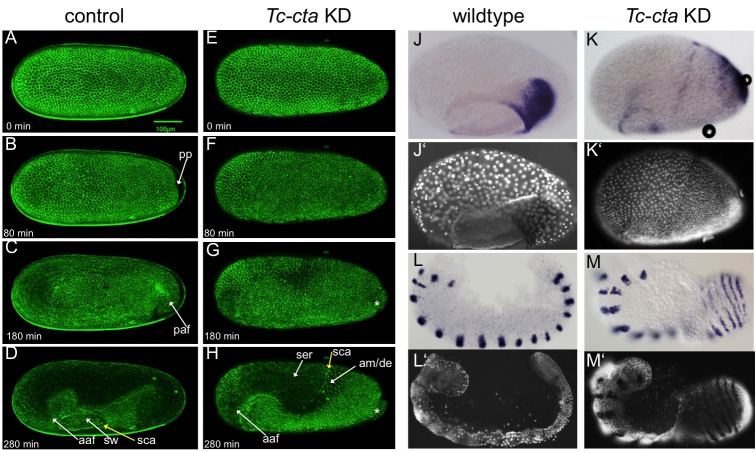
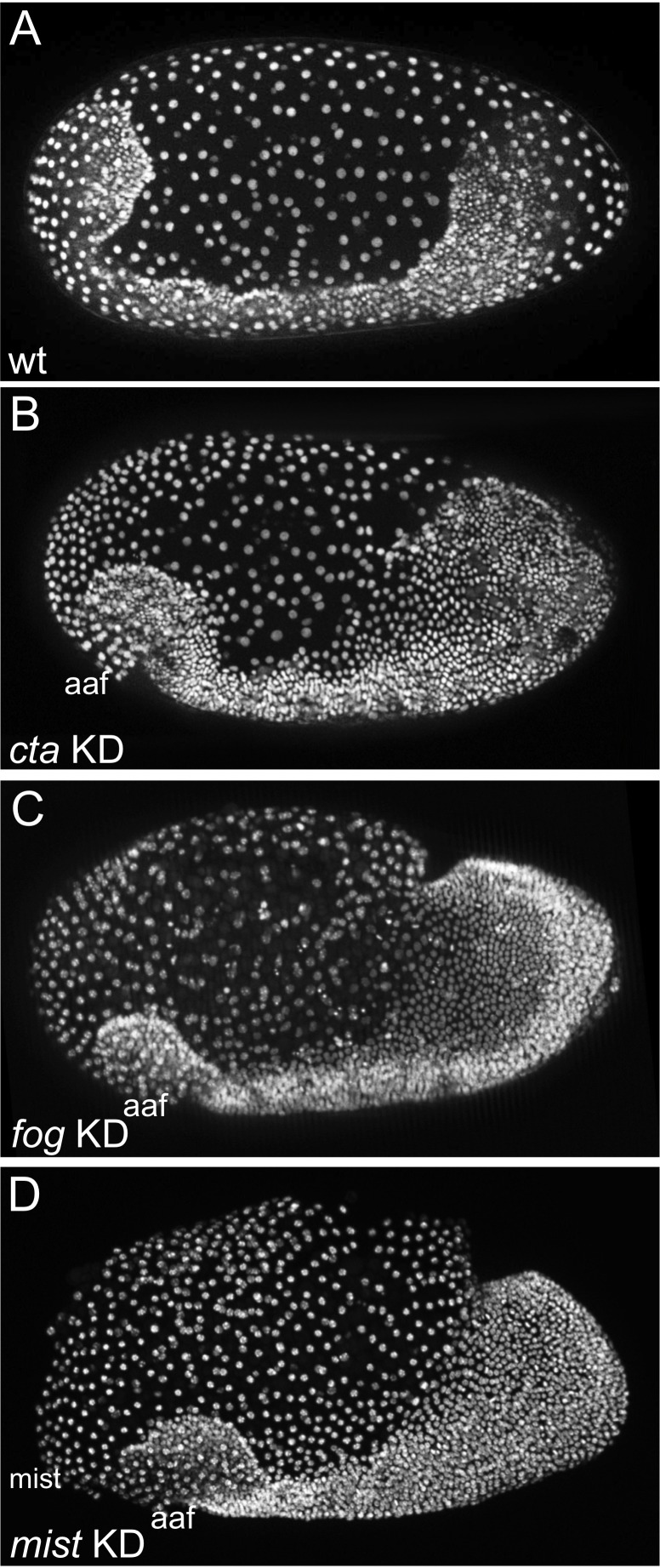
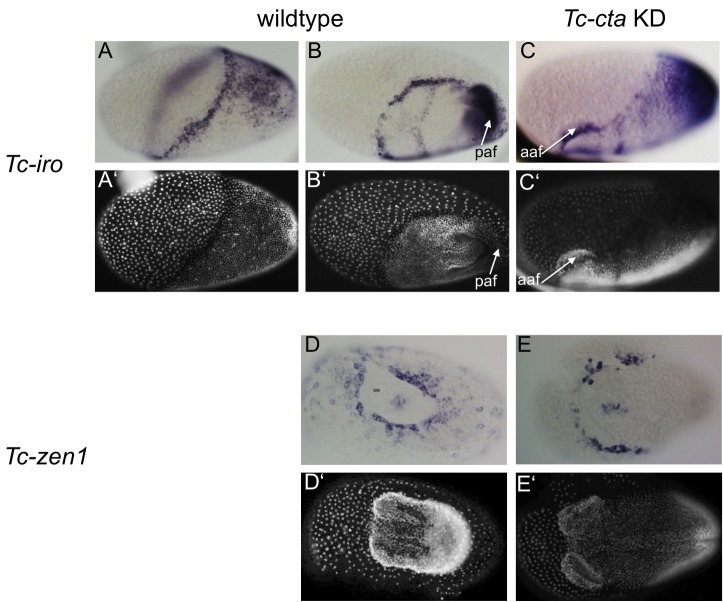
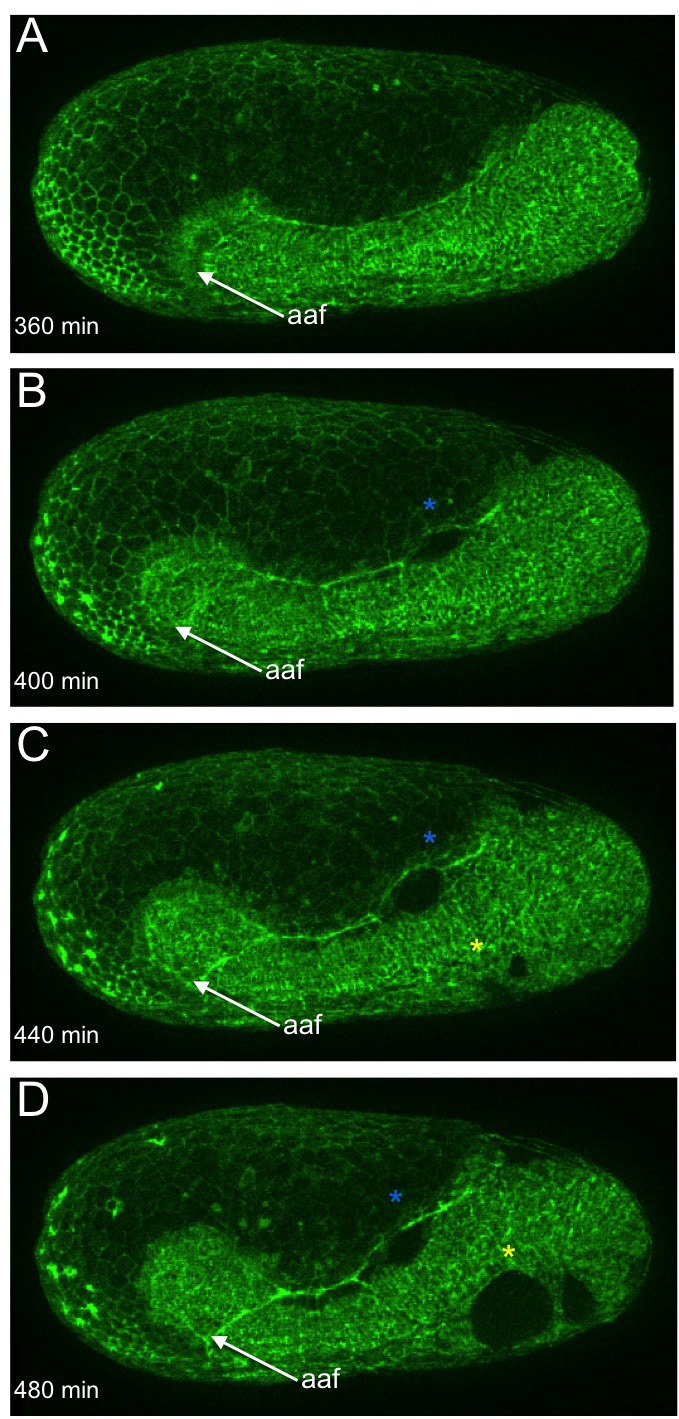
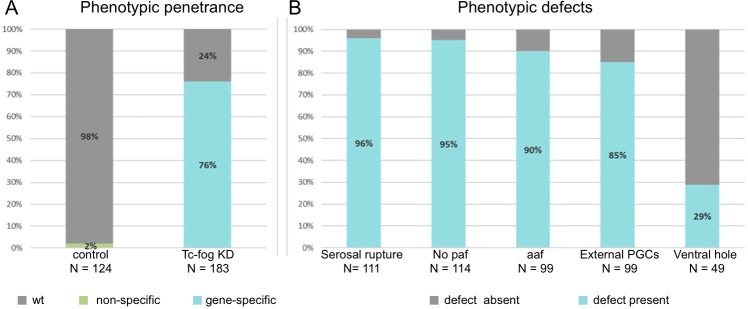
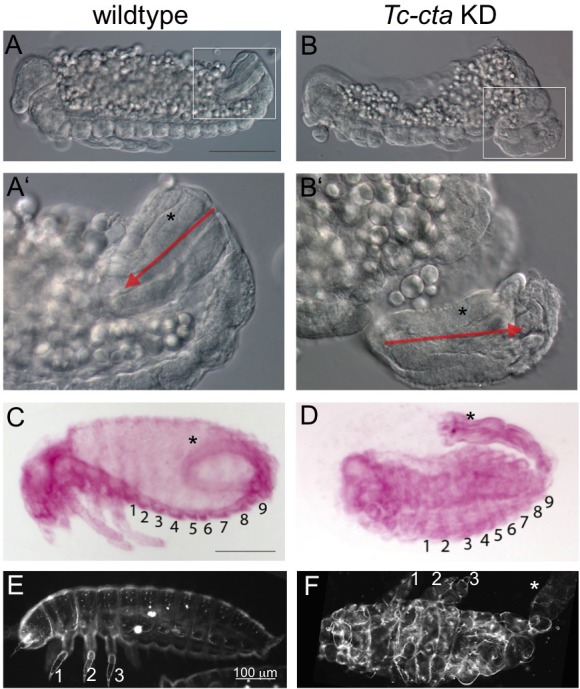
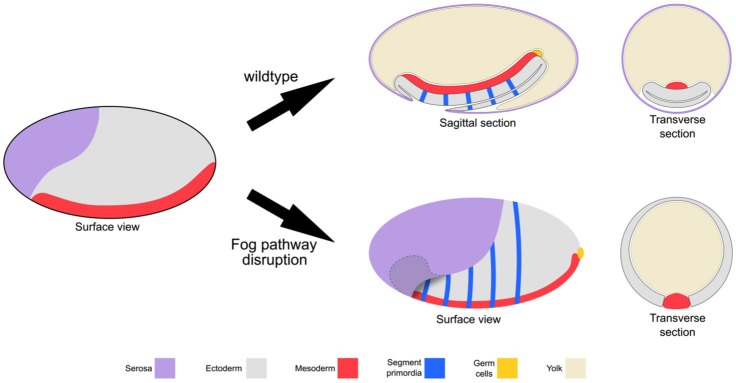
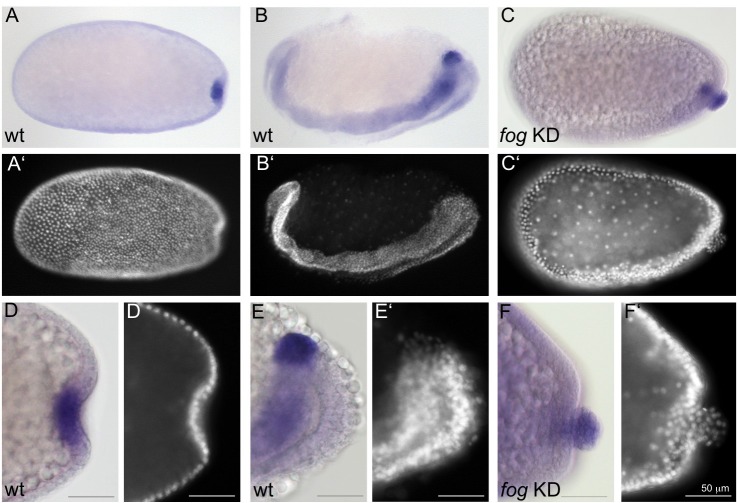
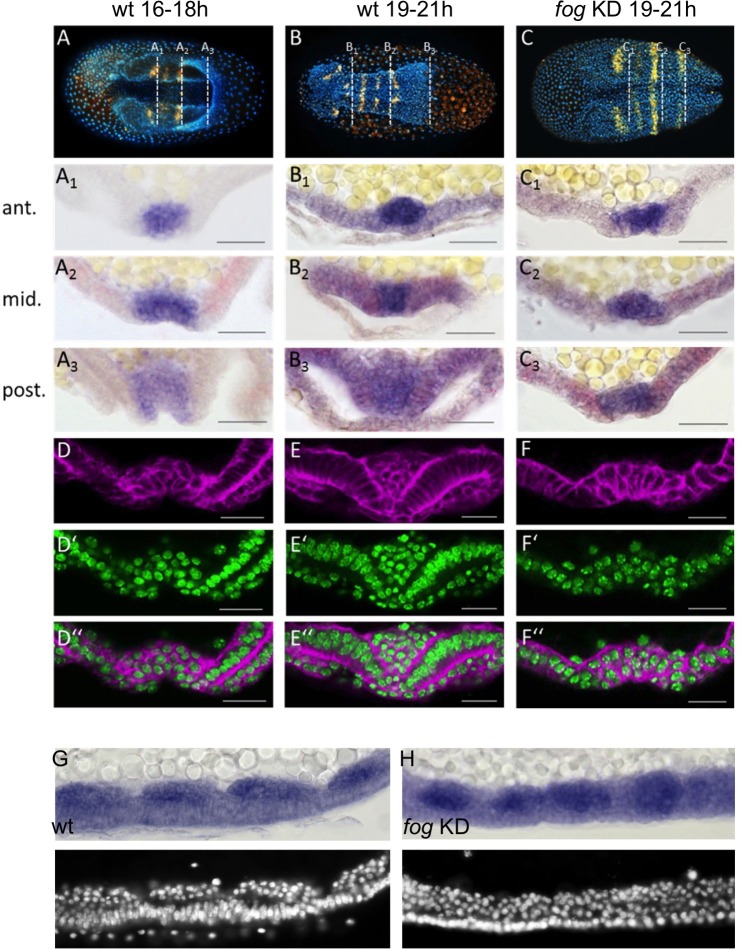
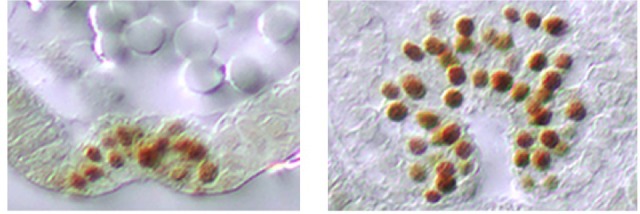
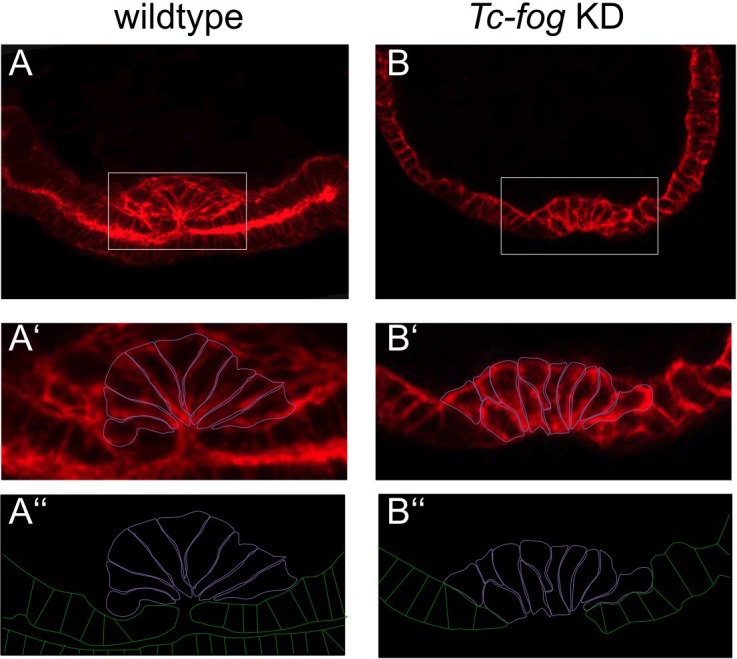
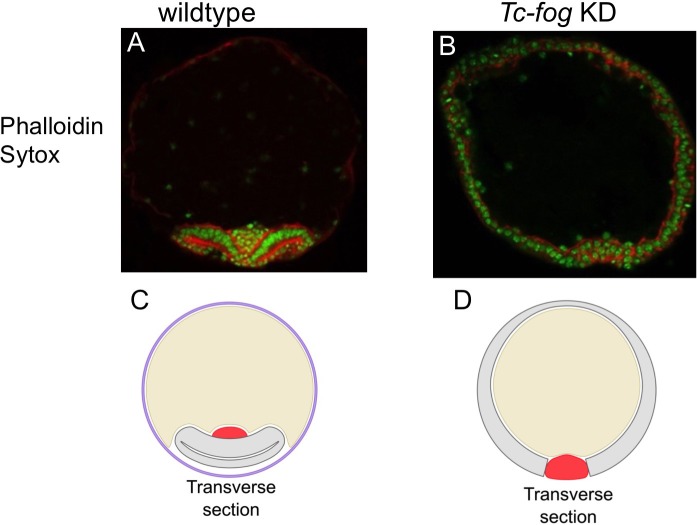
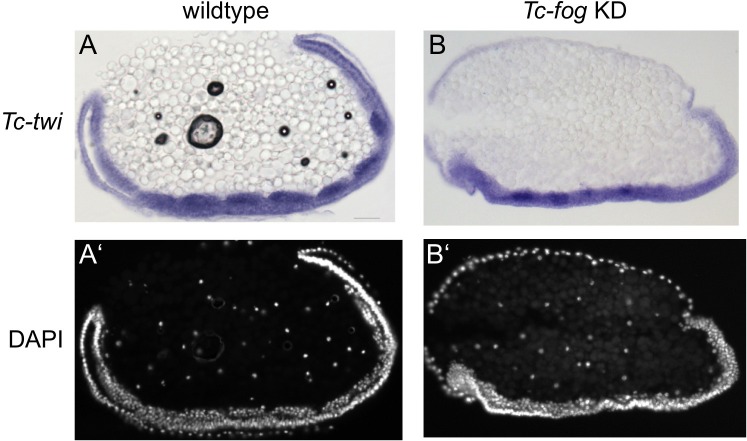
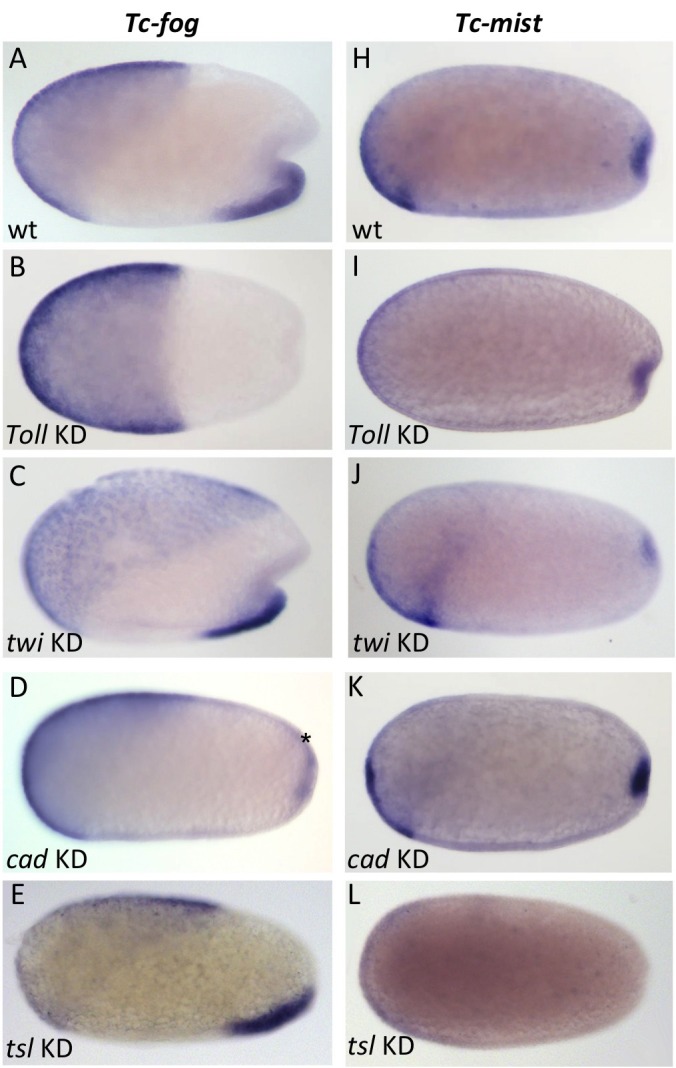
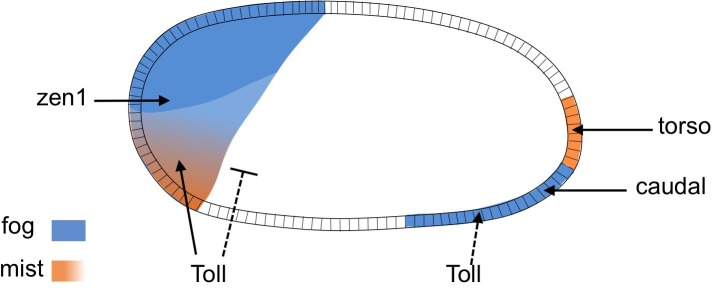
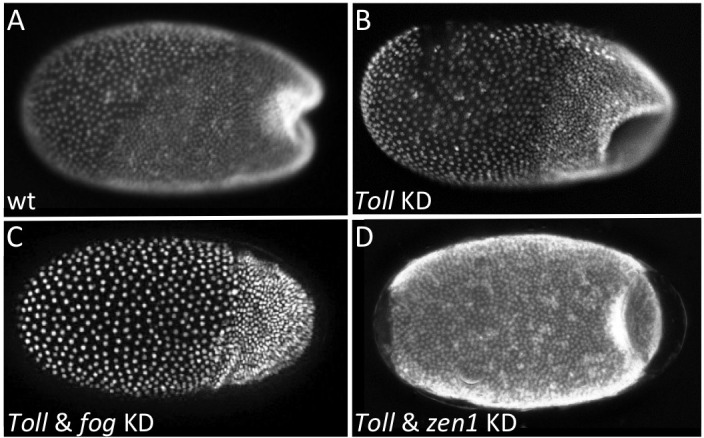
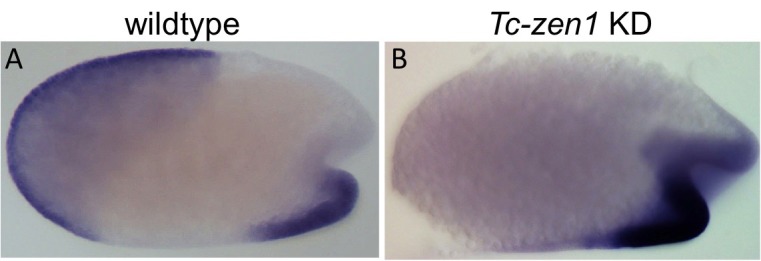
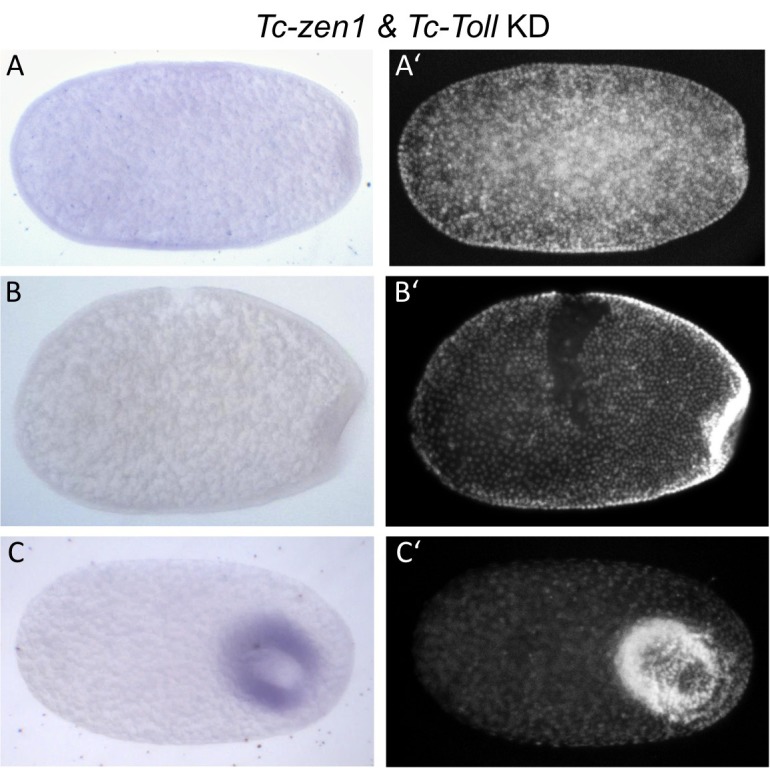
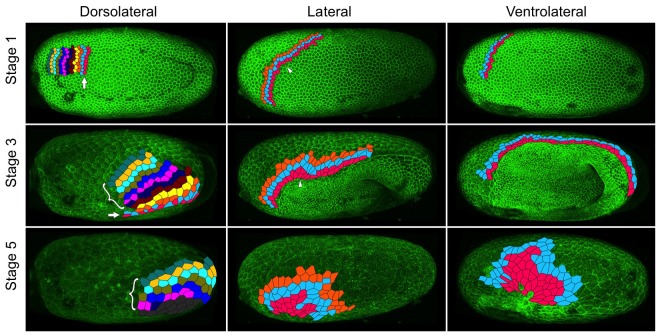
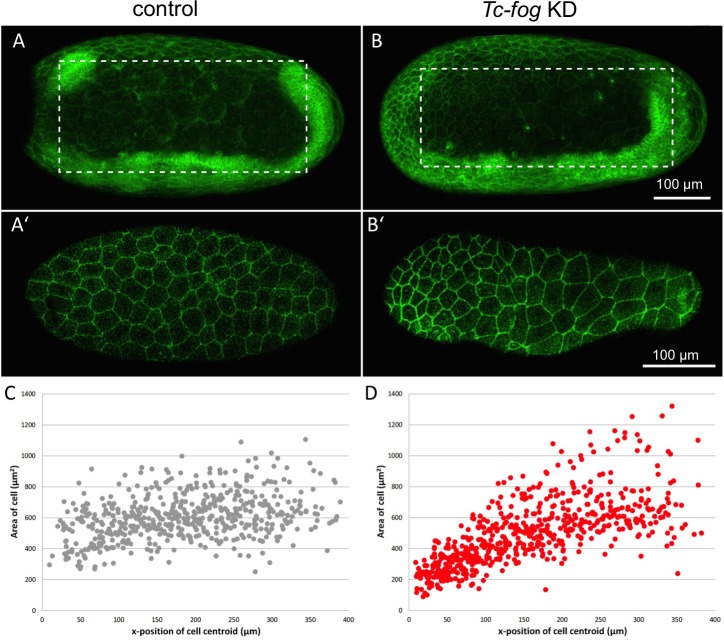
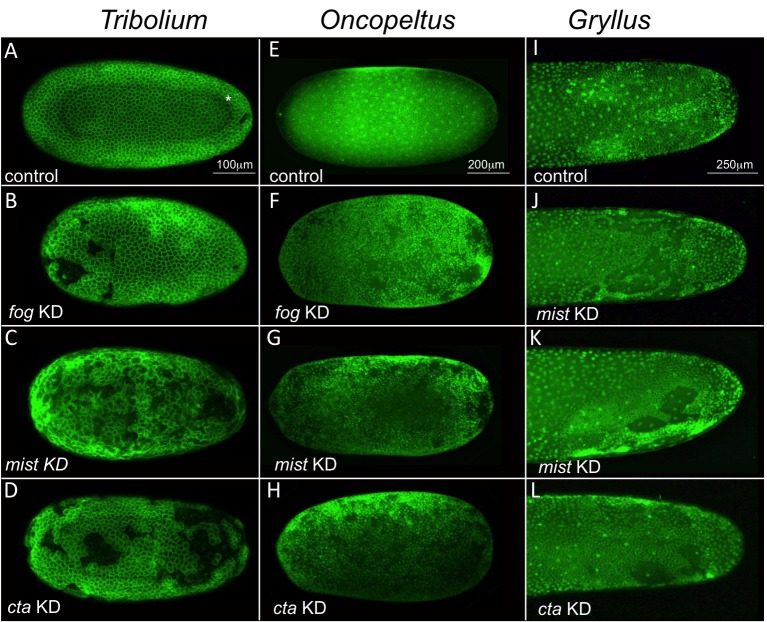
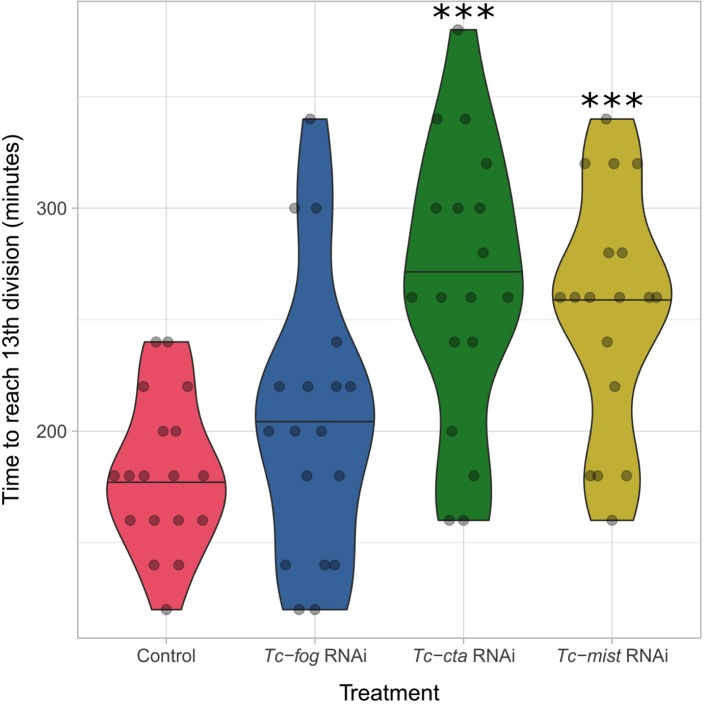
The Fog pathway represents one of the best-understood signaling cascades controlling epithelial morphogenesis. During gastrulation, Fog induces apical cell constrictions that drive the invagination of mesoderm and posterior gut primordia. The cellular mechanisms underlying primordia internalization vary greatly among insects and recent work has suggested that Fog signaling is specific to the fast mode of gastrulation found in some flies. On the contrary, here we show in the beetle , whose development is broadly representative for insects, that Fog has multiple morphogenetic functions. It modulates mesoderm internalization and controls a massive posterior infolding involved in gut and extraembryonic development. In addition, Fog signaling affects blastoderm cellularization, primordial germ cell positioning, and cuboidal-to-squamous cell shape transitions in the extraembryonic serosa. Comparative analyses with two other distantly related insect species reveals that Fog's role during cellularization is widely conserved and therefore might represent the ancestral function of the pathway.
Fog 通路是调控上皮形态发生的信号级联途径中研究得最为透彻的途径之一。在原肠胚形成过程中,Fog 诱导顶端细胞的缢缩,从而推动中胚层和后肠原基的内陷。原基内陷的细胞机制在不同的昆虫之间存在很大差异,最近的研究表明 Fog 信号通路是某些蝇类中快速原肠胚形成所特有的。相反,在这里我们在甲虫中发现,其发育广泛代表了昆虫,Fog 具有多种形态发生功能。它调节中胚层的内陷,并控制涉及肠和胚胎外发育的大规模后内折。此外,Fog 信号通路还影响胚胎细胞化、原始生殖细胞定位以及胚胎外浆膜的立方细胞到鳞状细胞的形状转变。与另外两个亲缘关系较远的昆虫物种的比较分析表明,Fog 在细胞化过程中的作用是广泛保守的,因此可能代表了该途径的祖先功能。