McArdle Laboratory for Cancer Research, Department of Oncology, University of Wisconsin at Madison, School of Medicine and Public Health, Madison, United States.
Biophysics program, University of Wisconsin at Madison, Madison, United States.
Elife. 2022 Aug 4;11:e79736. doi: 10.7554/eLife.79736.
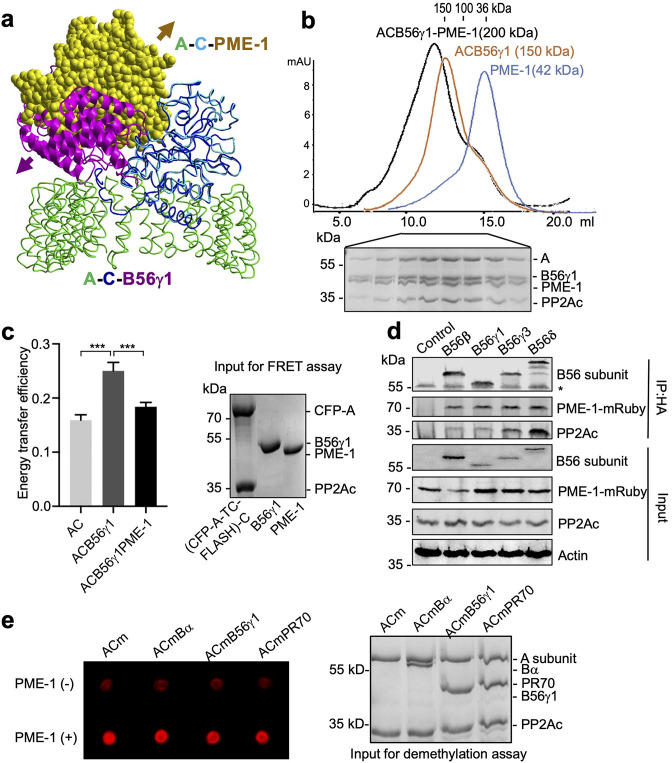
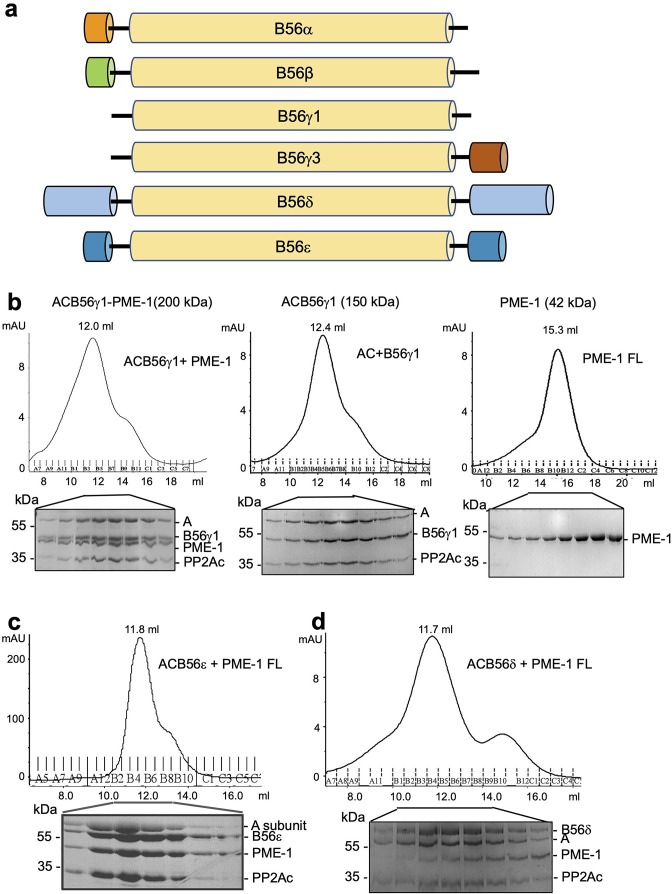
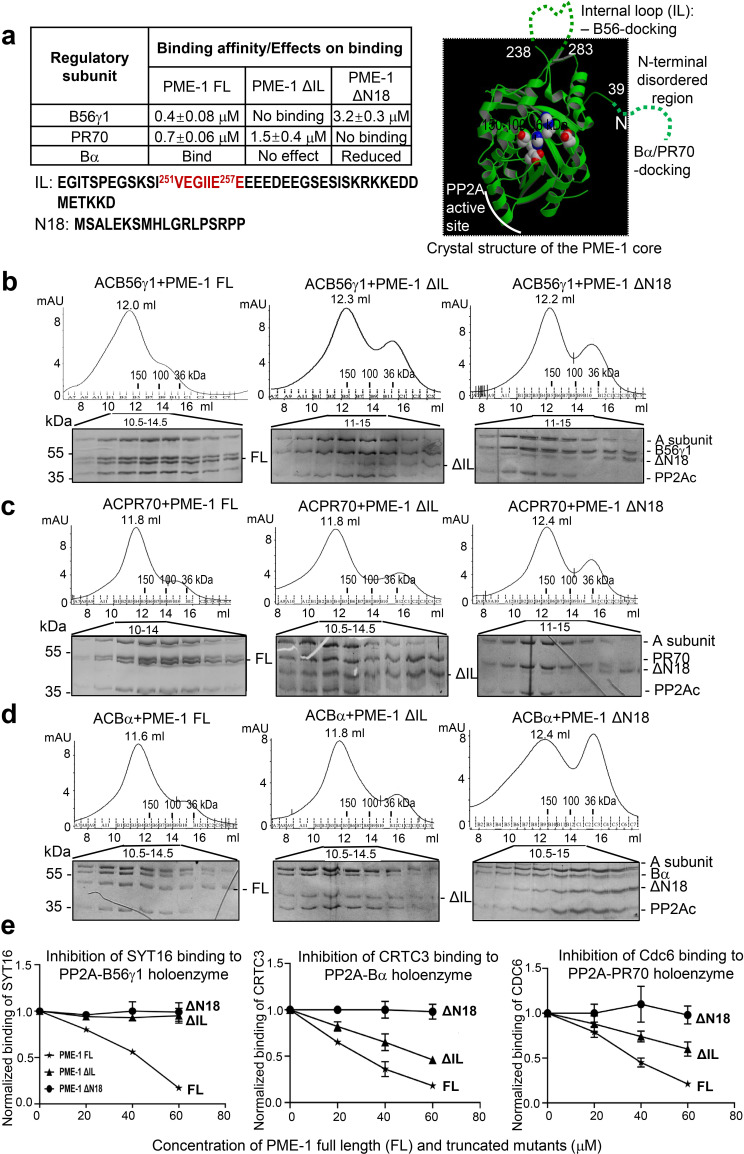
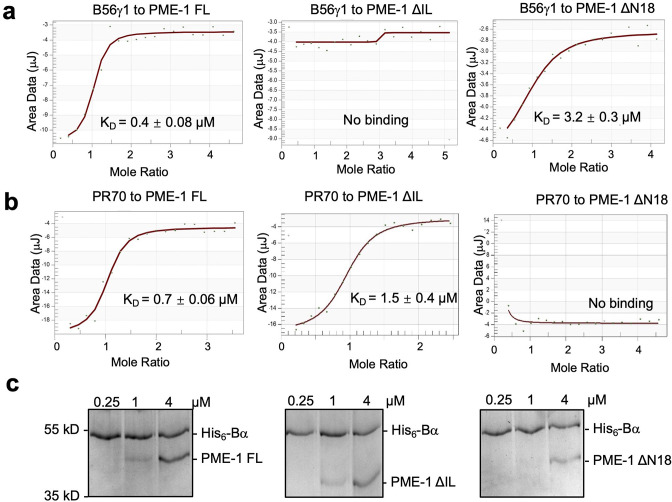
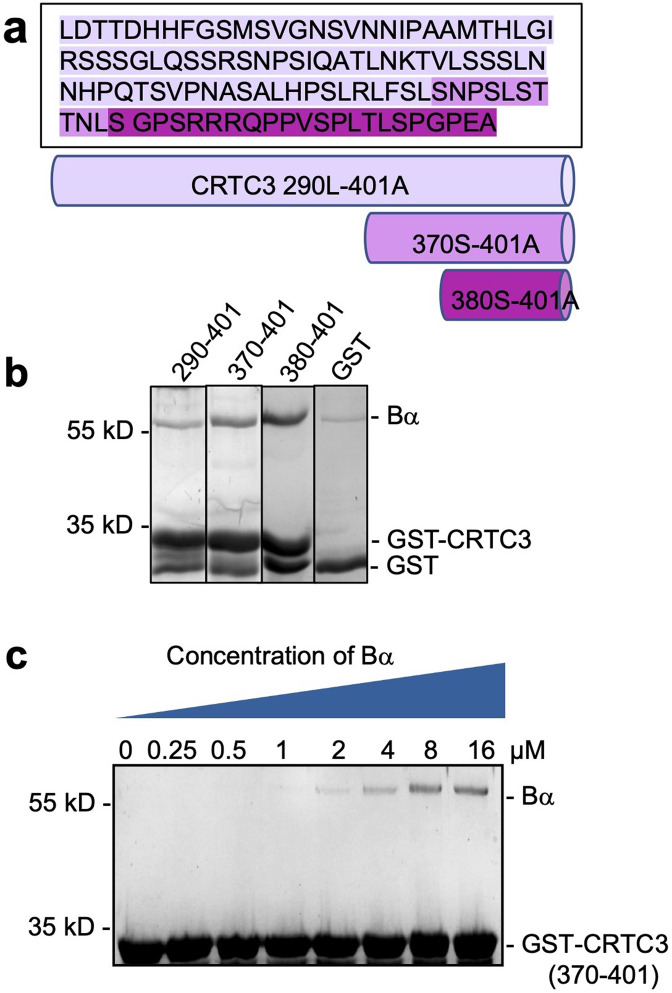
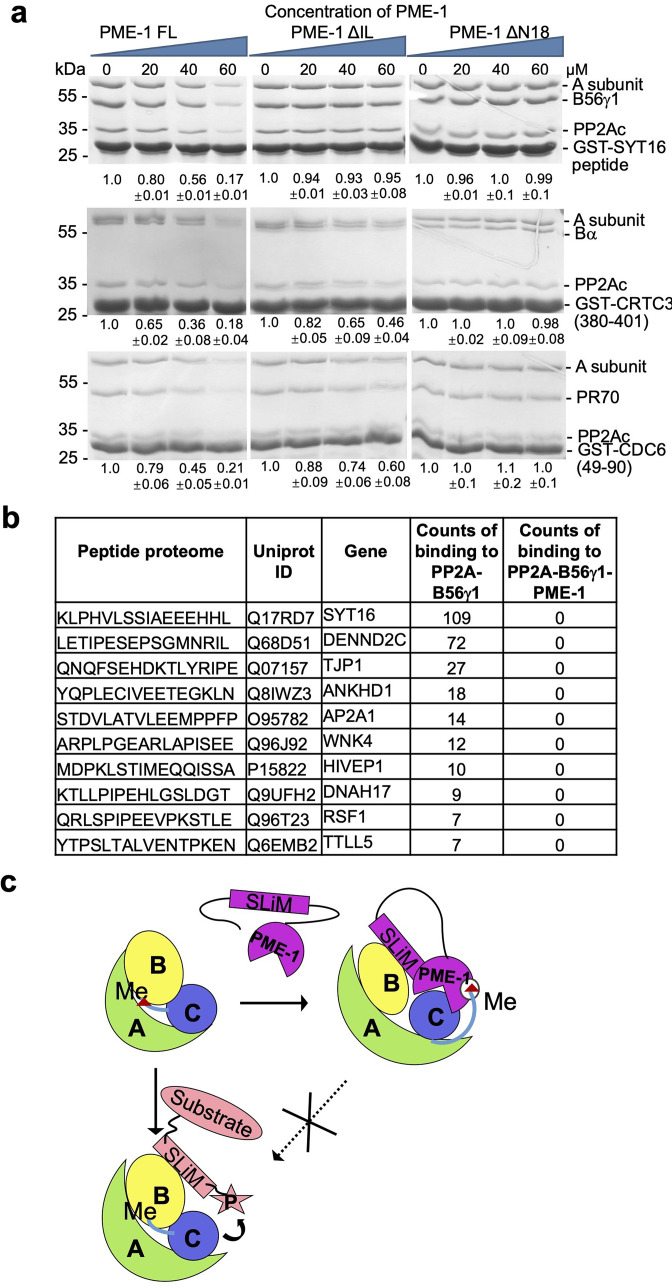
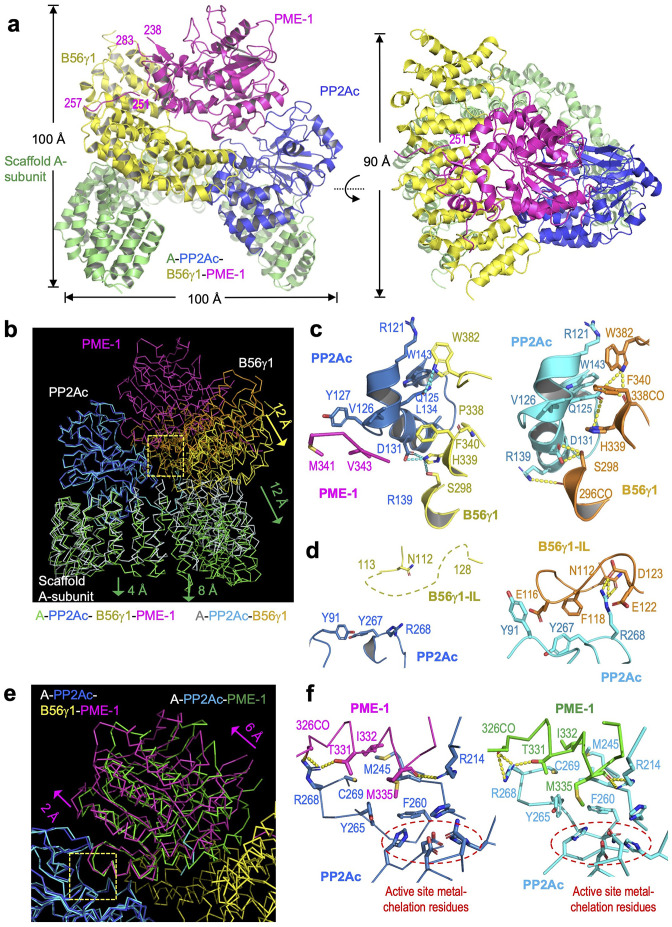
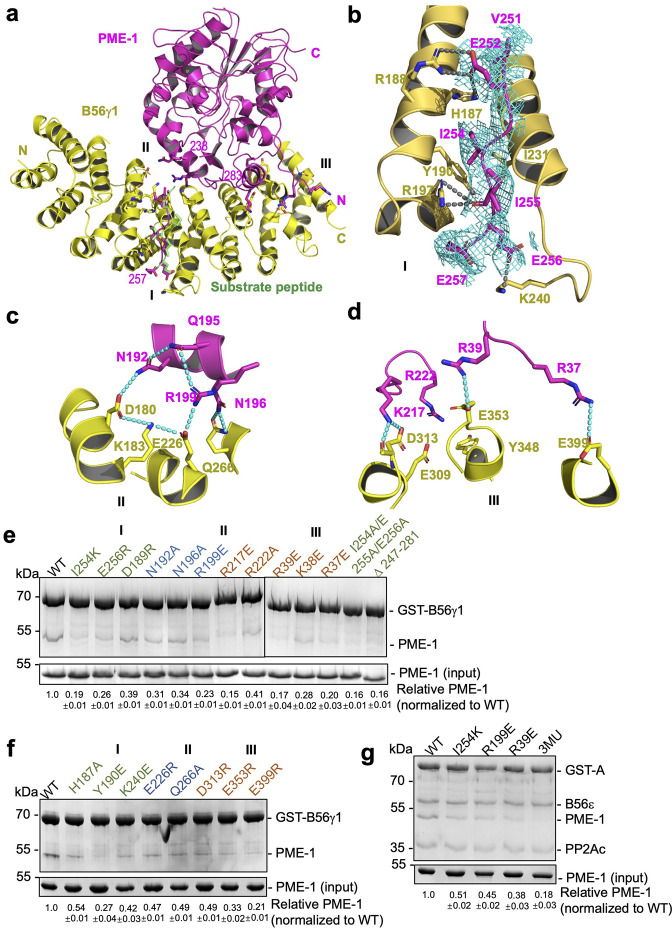
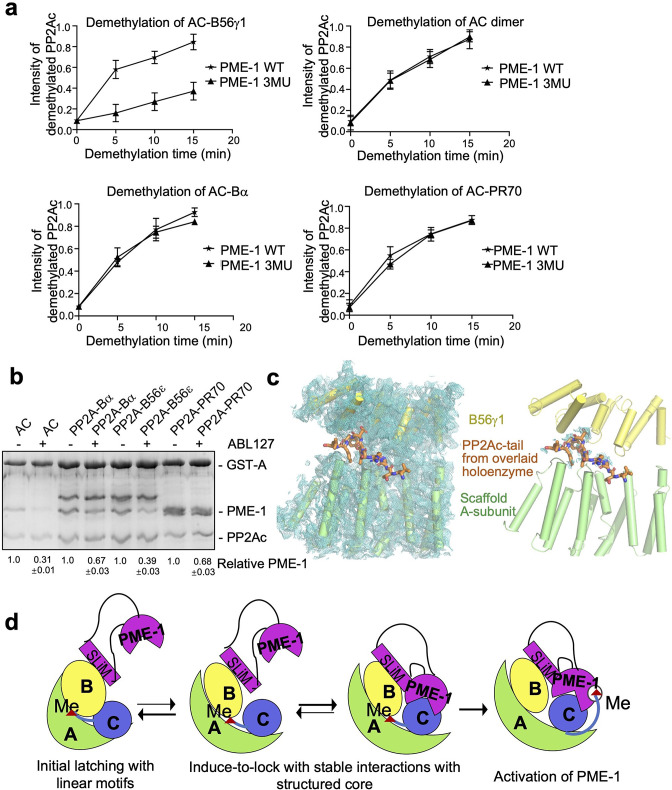
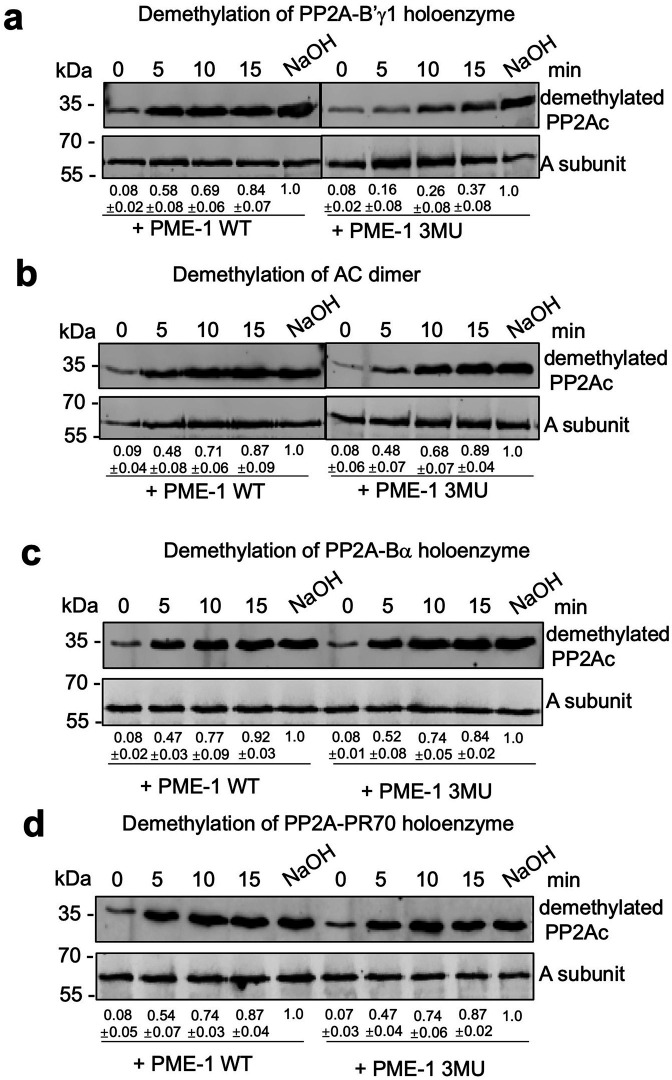
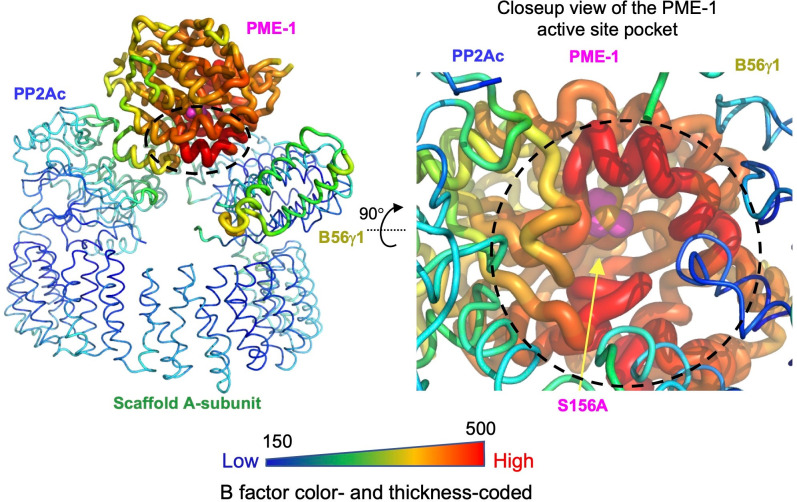
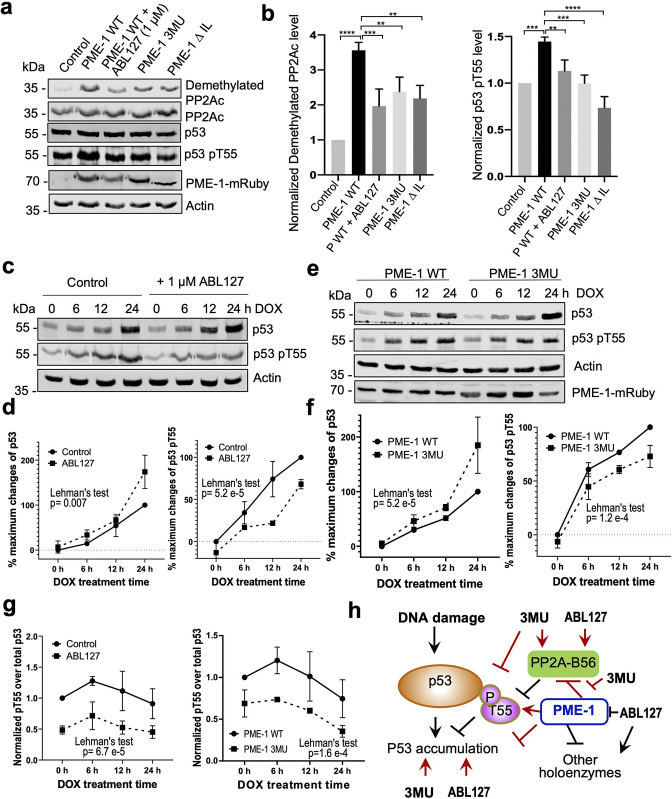
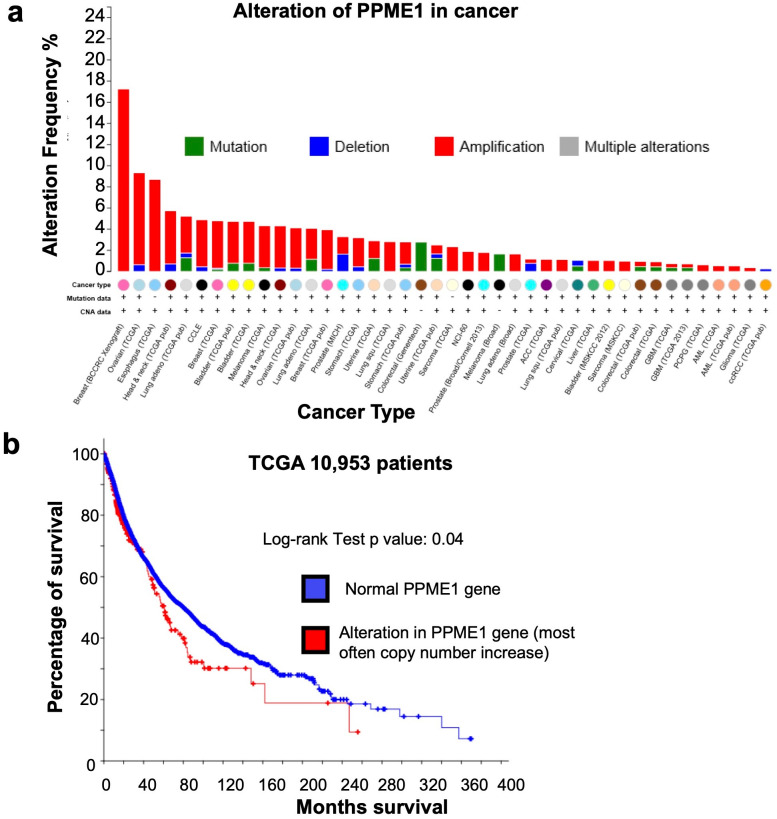
Protein phosphatase 2A (PP2A) holoenzymes target broad substrates by recognizing short motifs via regulatory subunits. PP2A methylesterase 1 (PME-1) is a cancer-promoting enzyme and undergoes methylesterase activation upon binding to the PP2A core enzyme. Here, we showed that PME-1 readily demethylates different families of PP2A holoenzymes and blocks substrate recognition in vitro. The high-resolution cryoelectron microscopy structure of a PP2A-B56 holoenzyme-PME-1 complex reveals that PME-1 disordered regions, including a substrate-mimicking motif, tether to the B56 regulatory subunit at remote sites. They occupy the holoenzyme substrate-binding groove and allow large structural shifts in both holoenzyme and PME-1 to enable multipartite contacts at structured cores to activate the methylesterase. B56 interface mutations selectively block PME-1 activity toward PP2A-B56 holoenzymes and affect the methylation of a fraction of total cellular PP2A. The B56 interface mutations allow us to uncover B56-specific PME-1 functions in p53 signaling. Our studies reveal multiple mechanisms of PME-1 in suppressing holoenzyme functions and versatile PME-1 activities derived from coupling substrate-mimicking motifs to dynamic structured cores.
蛋白磷酸酶 2A(PP2A)全酶通过调节亚基识别短序列来靶向广泛的底物。PP2A 甲酯酶 1(PME-1)是一种促进癌症的酶,与 PP2A 核心酶结合后会发生甲酯酶激活。在这里,我们表明 PME-1 可以轻易地去甲基化不同家族的 PP2A 全酶,并在体外阻断底物识别。PP2A-B56 全酶-PME-1 复合物的高分辨率冷冻电子显微镜结构表明,PME-1 的无规则区域,包括一个底物模拟序列,通过远程位点与 B56 调节亚基连接。它们占据全酶的底物结合槽,并允许全酶和 PME-1 发生大的结构位移,从而在结构化核心中实现多部分接触,以激活甲酯酶。B56 界面突变选择性地阻止 PME-1 对 PP2A-B56 全酶的活性,并影响总细胞 PP2A 的一部分甲基化。B56 界面突变使我们能够揭示 B56 特异性 PME-1 在 p53 信号转导中的功能。我们的研究揭示了 PME-1 抑制全酶功能的多种机制,以及源自将底物模拟序列与动态结构化核心结合的多功能 PME-1 活性。