Department of Biochemistry and Molecular Biology, Faculty of Medicine, University of Debrecen, Hungary; Doctoral School of Molecular Cell and Immune Biology, University of Debrecen, Debrecen, Hungary.
Basic Science Program, Frederick National Laboratory for Cancer Research, Frederick, MD 21702, USA.
J Mol Biol. 2023 Mar 15;435(6):168012. doi: 10.1016/j.jmb.2023.168012. Epub 2023 Feb 13.
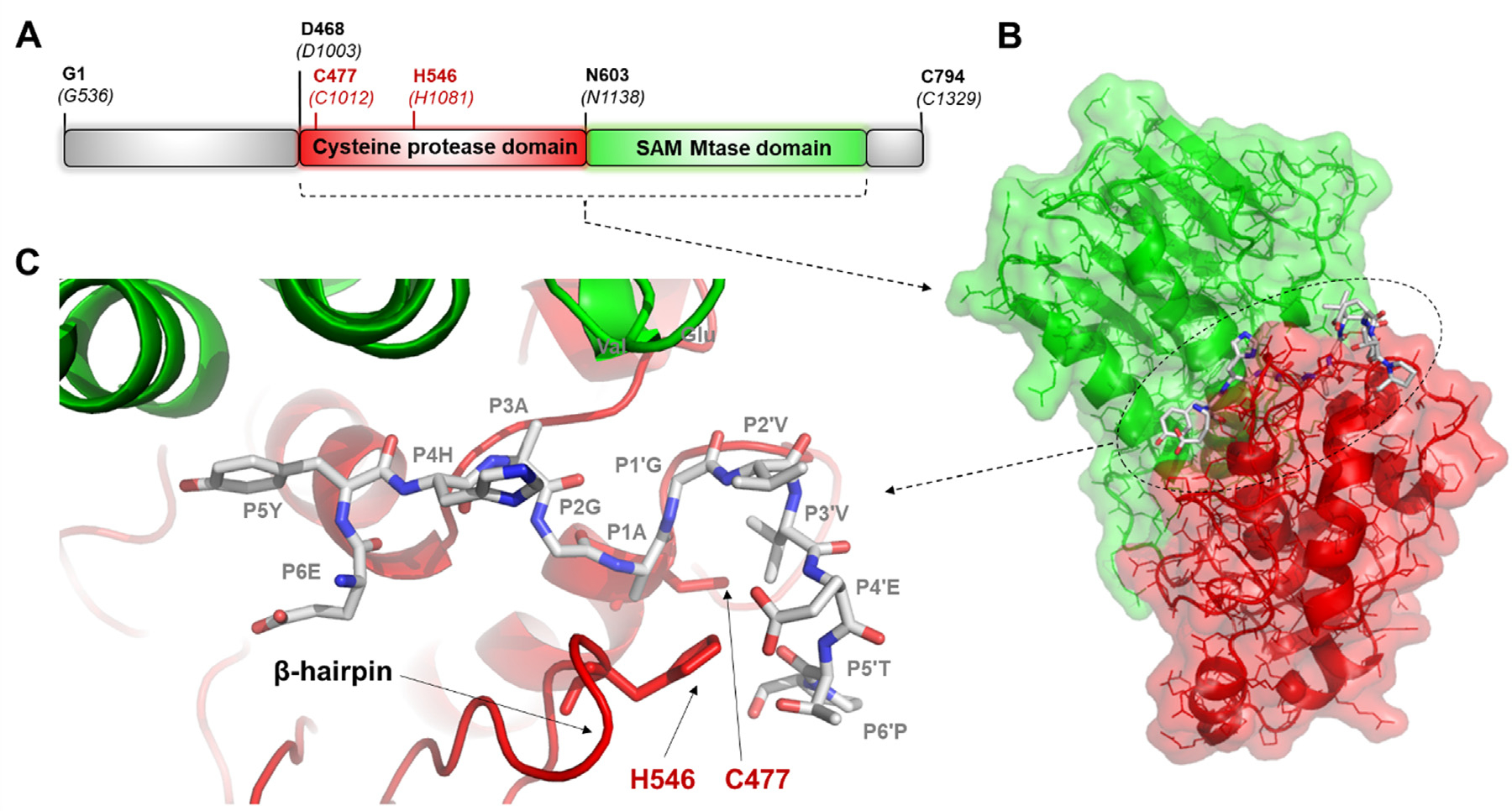
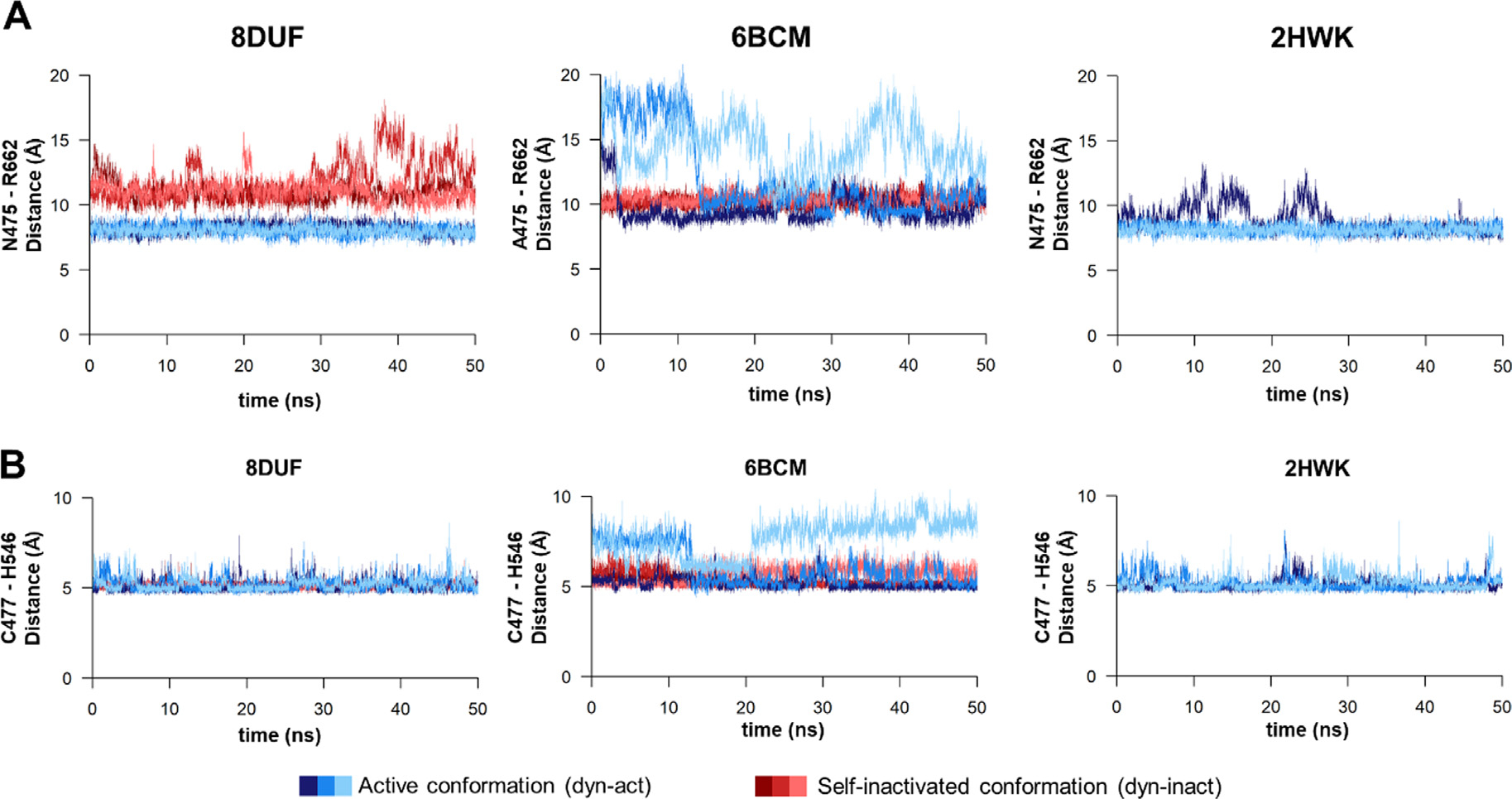
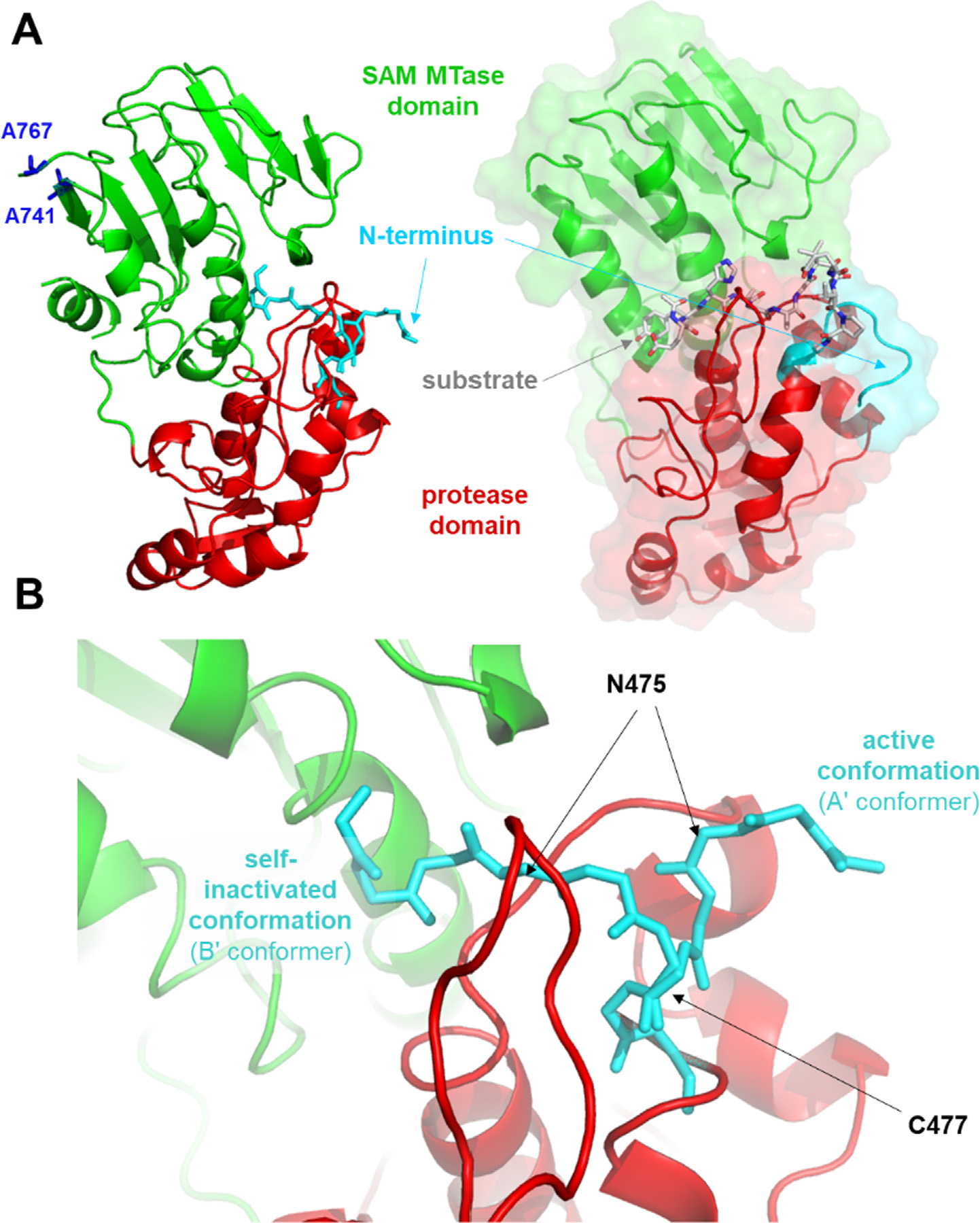
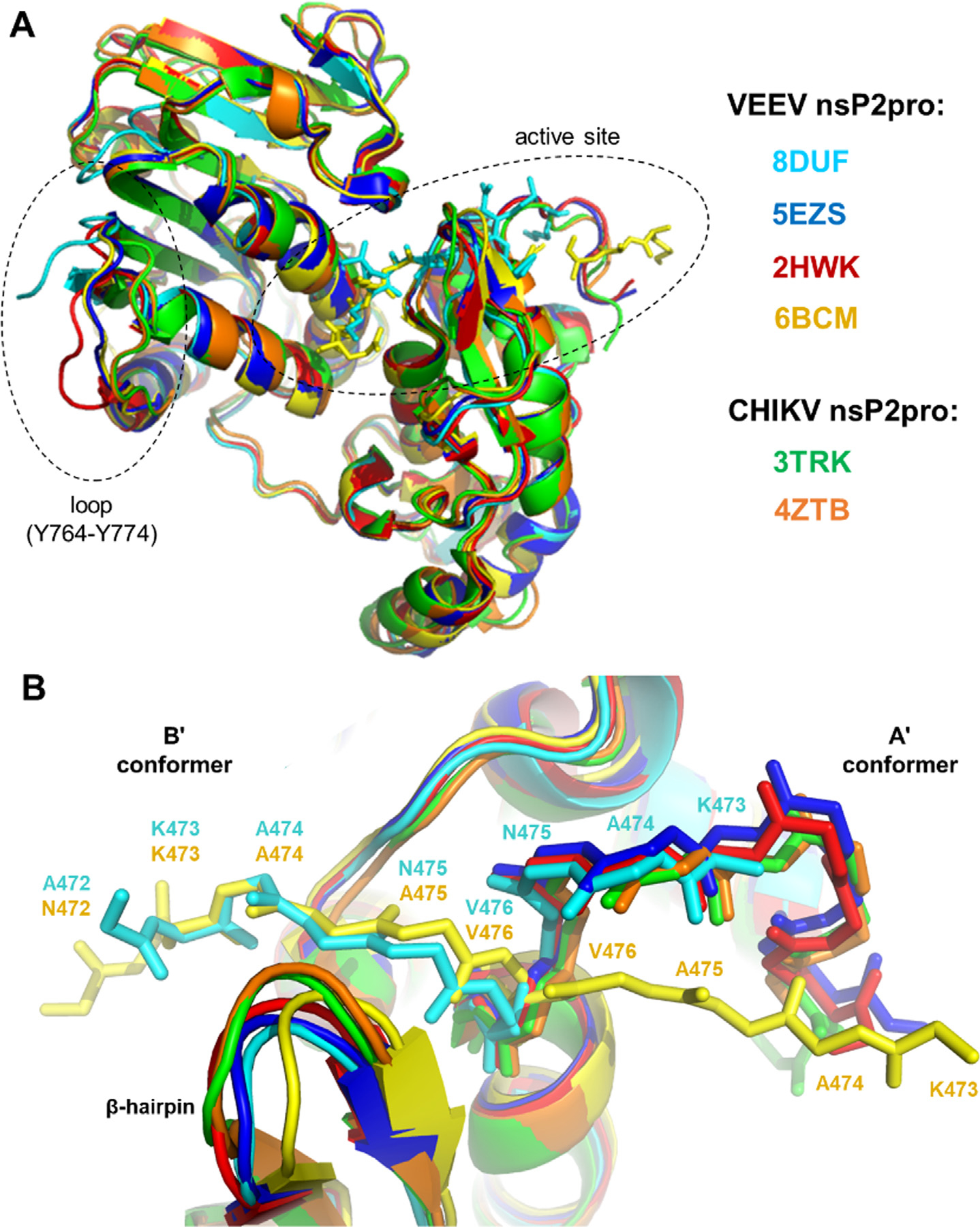
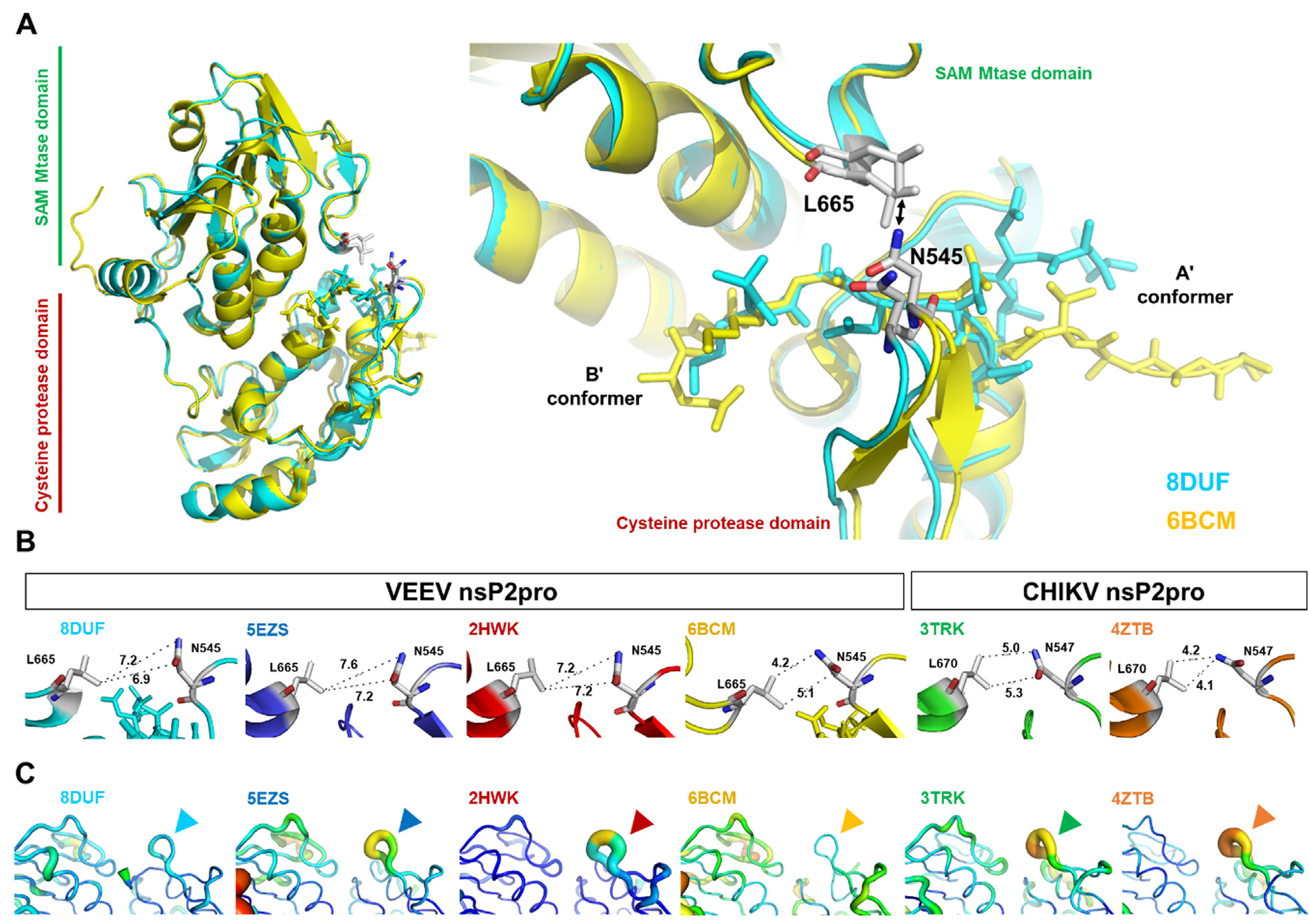
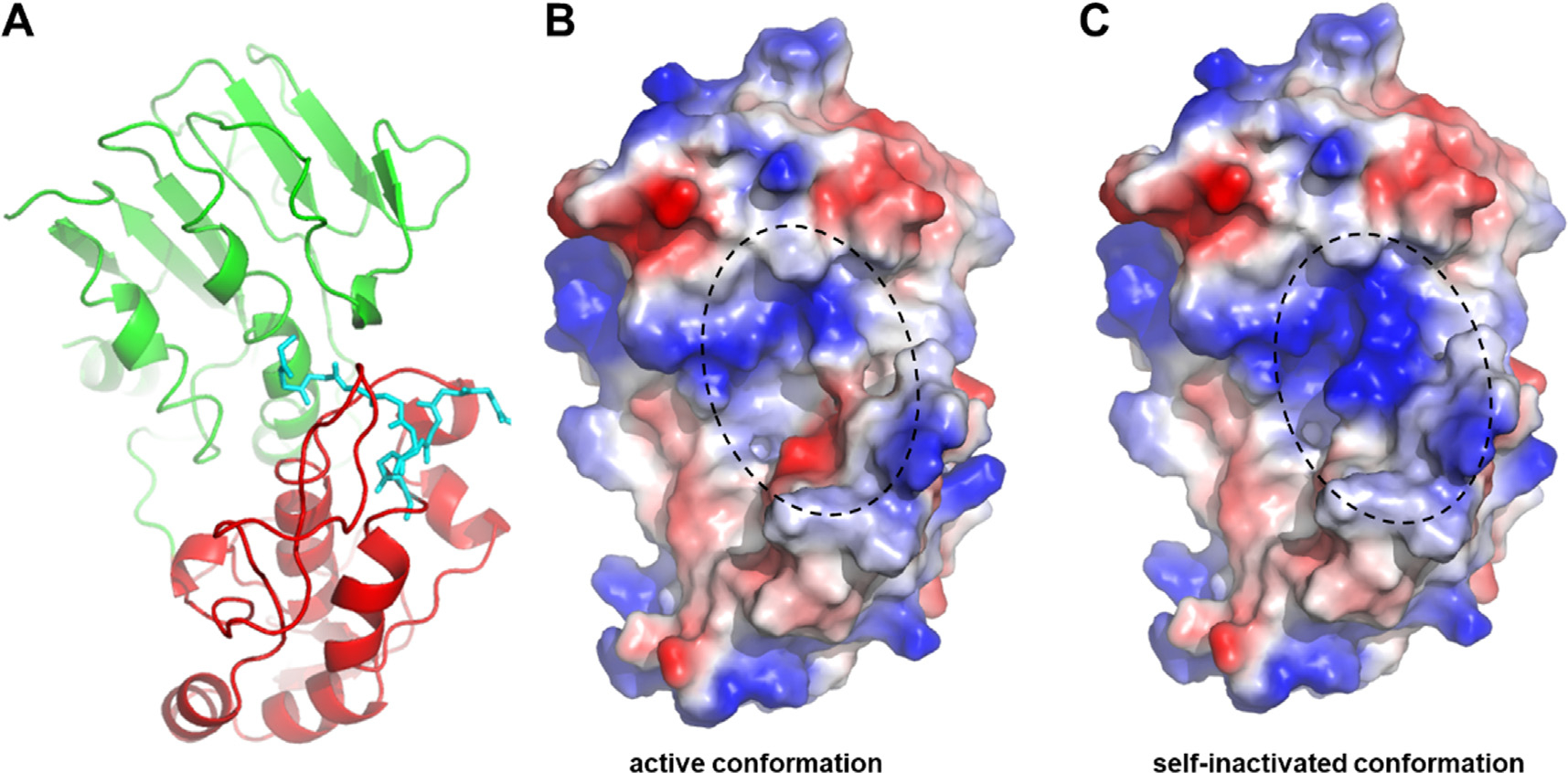
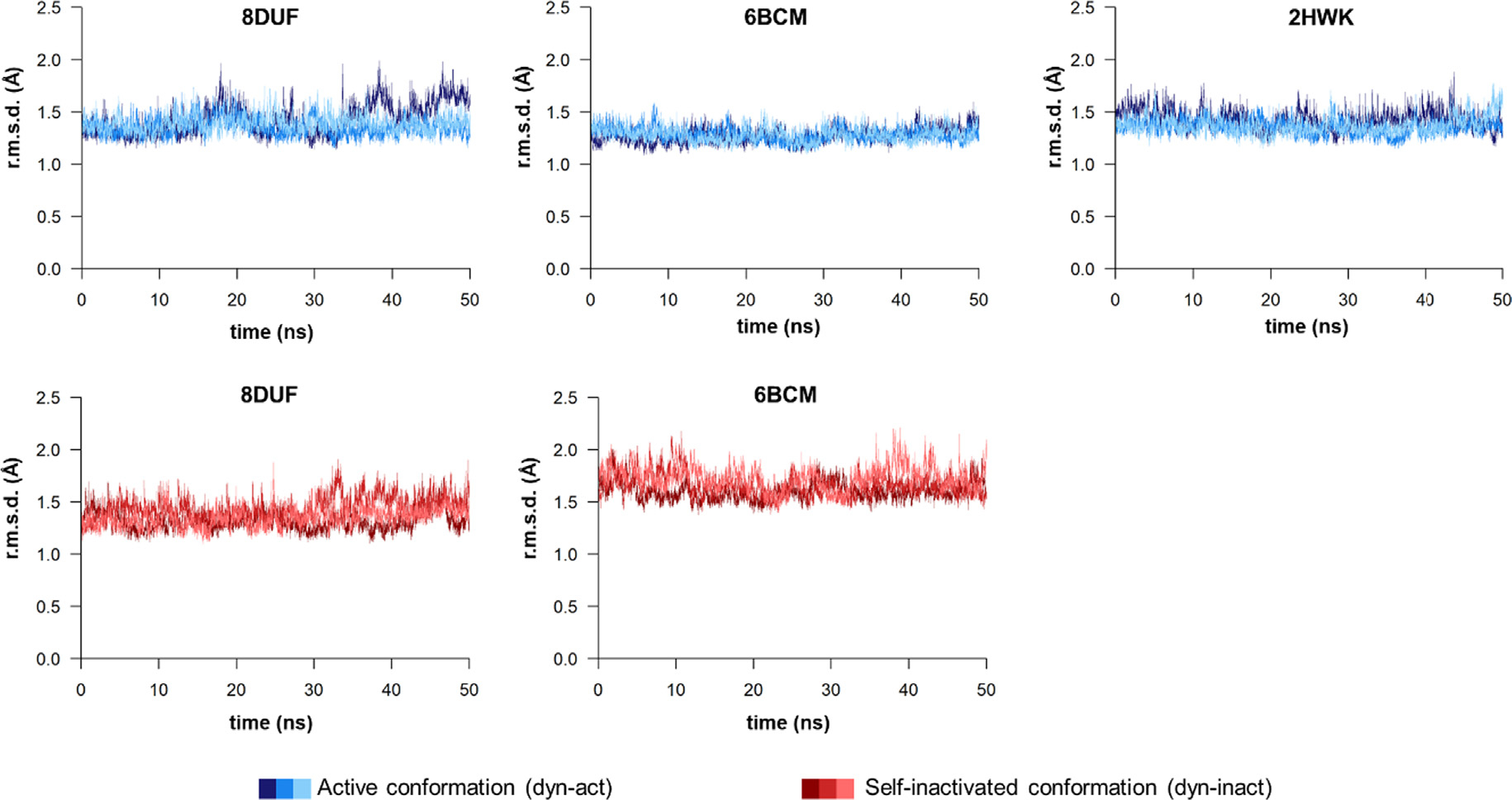
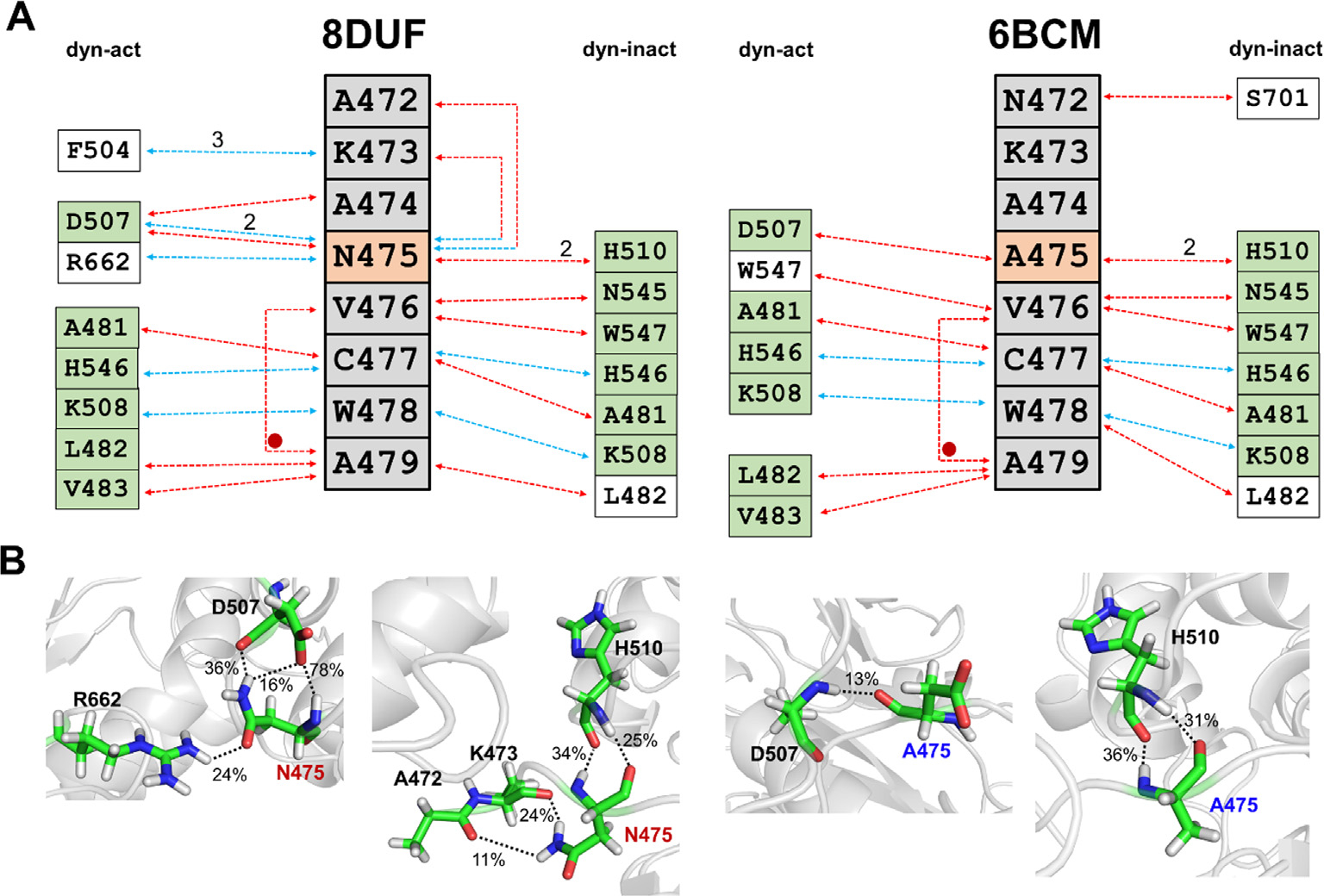
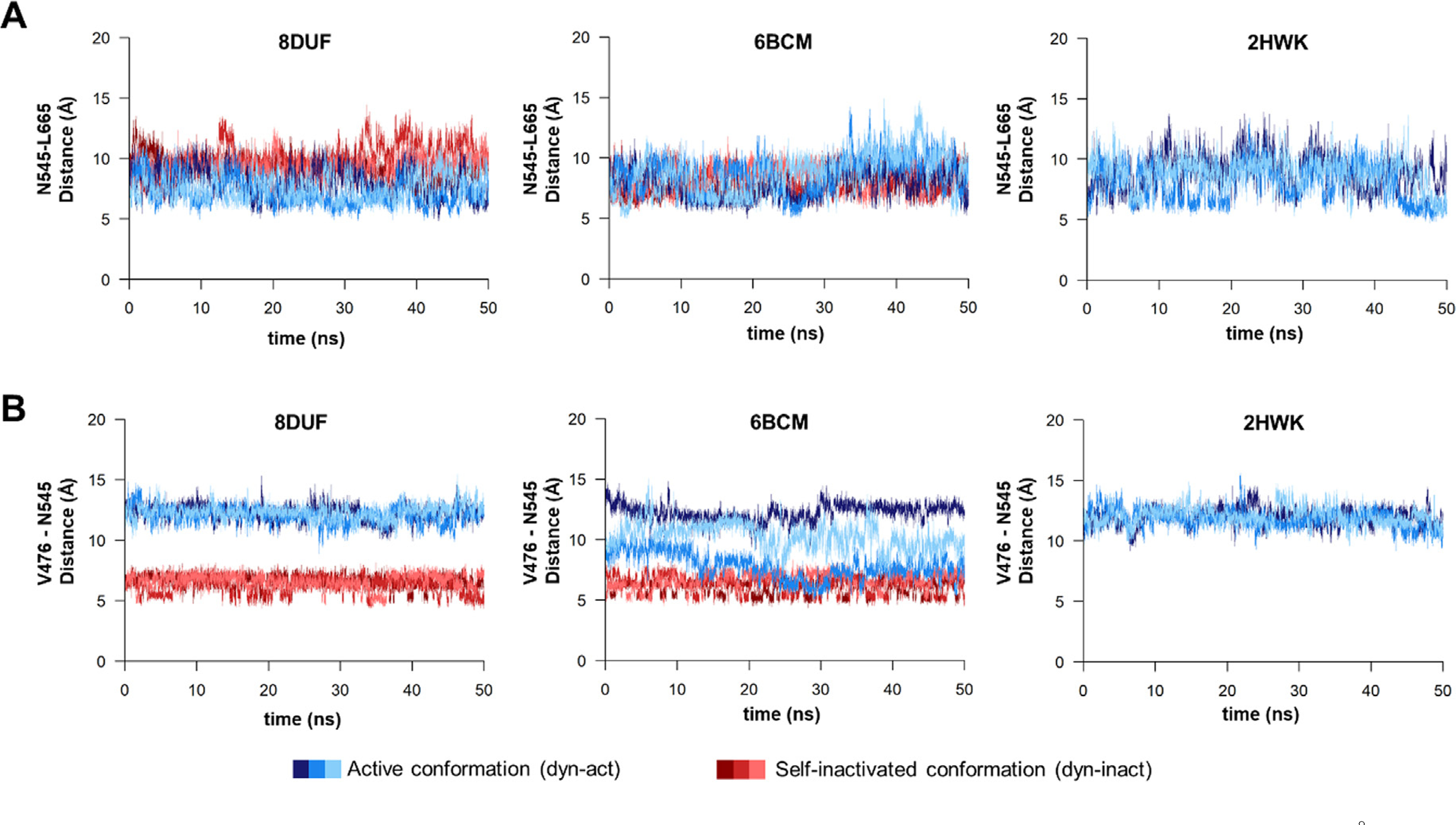
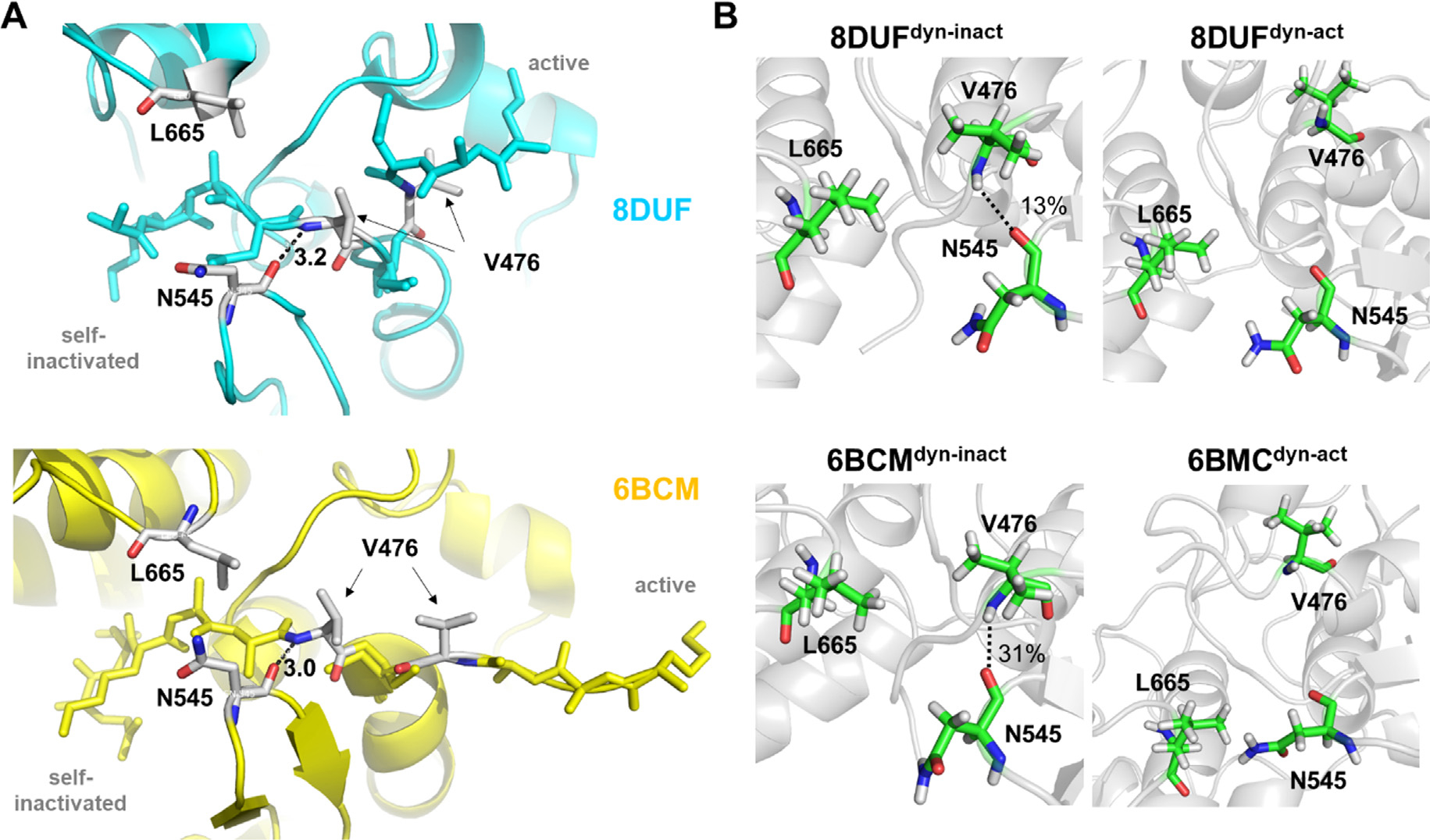
The Venezuelan equine encephalitis virus (VEEV) belongs to the Togaviridae family and is pathogenic to both humans and equines. The VEEV non-structural protein 2 (nsP2) is a cysteine protease (nsP2pro) that processes the polyprotein and thus it is a drug target for inhibitor discovery. The atomic structure of the VEEV nsP2 catalytic domain was previously characterized by both X-ray crystallography and computational studies. A modified nsP2pro harboring a N475A mutation in the N terminus was observed to exhibit an unexpected conformation: the N-terminal residues bind to the active site, mimicking binding of a substrate. The large conformational change of the N terminus was assumed to be induced by the N475A mutation, as N475 has an important role in stabilization of the N terminus and the active site. This conformation was first observed in the N475A mutant, but we also found it while determining a crystal structure of the catalytically active nsP2pro containing the wild-type N475 active site residue and K741A/K767A surface entropy reduction mutations. This suggests that the N475A mutation is not a prerequisite for self-inhibition. Here, we describe a high resolution (1.46 Å) crystal structure of a truncated nsP2pro (residues 463-785, K741A/K767A) and analyze the structure further by molecular dynamics to study the active and self-inhibited conformations of nsP2pro and its N475A mutant. A comparison of the different conformations of the N-terminal residues sheds a light on the interactions that play an important role in the stabilization of the enzyme.
委内瑞拉马脑炎病毒(VEEV)属于披膜病毒科,对人类和马都具有致病性。VEEV 的非结构蛋白 2(nsP2)是一种半胱氨酸蛋白酶(nsP2pro),可对多蛋白进行加工,因此是抑制剂发现的药物靶点。VEEV nsP2 催化结构域的原子结构先前已通过 X 射线晶体学和计算研究进行了表征。观察到具有 N 端 N475A 突变的修饰 nsP2pro 表现出出乎意料的构象:N 端残基结合到活性部位,模拟底物结合。假定 N 端的大构象变化是由 N475A 突变引起的,因为 N475 在 N 端和活性部位的稳定中起重要作用。这种构象首先在 N475A 突变体中观察到,但我们在确定包含野生型 N475 活性部位残基和 K741A/K767A 表面熵降低突变的催化活性 nsP2pro 的晶体结构时也发现了它。这表明 N475A 突变不是自我抑制的先决条件。在这里,我们描述了一个截断的 nsP2pro(残基 463-785,K741A/K767A)的高分辨率(1.46 Å)晶体结构,并通过分子动力学进一步分析该结构,以研究 nsP2pro 及其 N475A 突变体的活性和自我抑制构象。比较 N 端残基的不同构象揭示了在酶稳定中起重要作用的相互作用。