Department of Pediatrics, Columbia University Vagelos College of Physicians and Surgeons, New York, New York, USA.
Center for Host-Pathogen Interaction, Columbia University Vagelos College of Physicians and Surgeons, New York, New York, USA.
mBio. 2024 Nov 13;15(11):e0232724. doi: 10.1128/mbio.02327-24. Epub 2024 Oct 9.
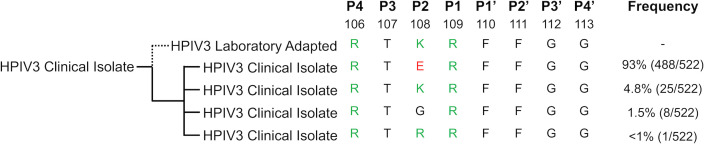
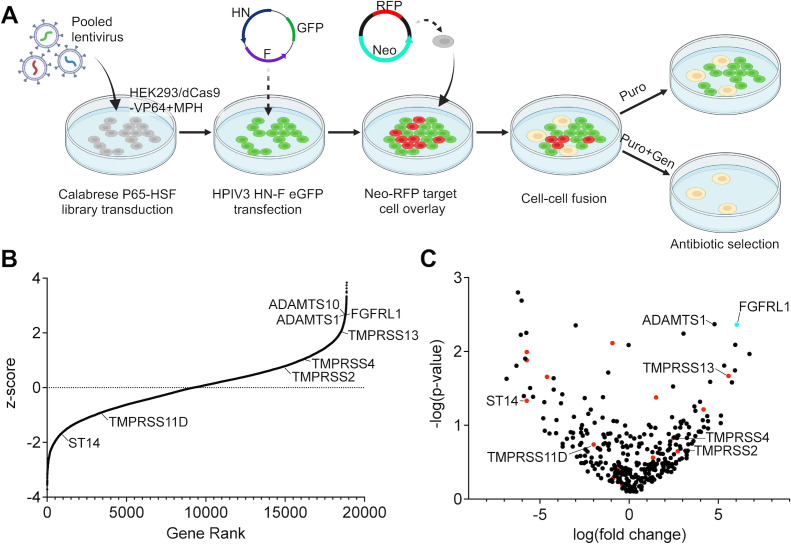
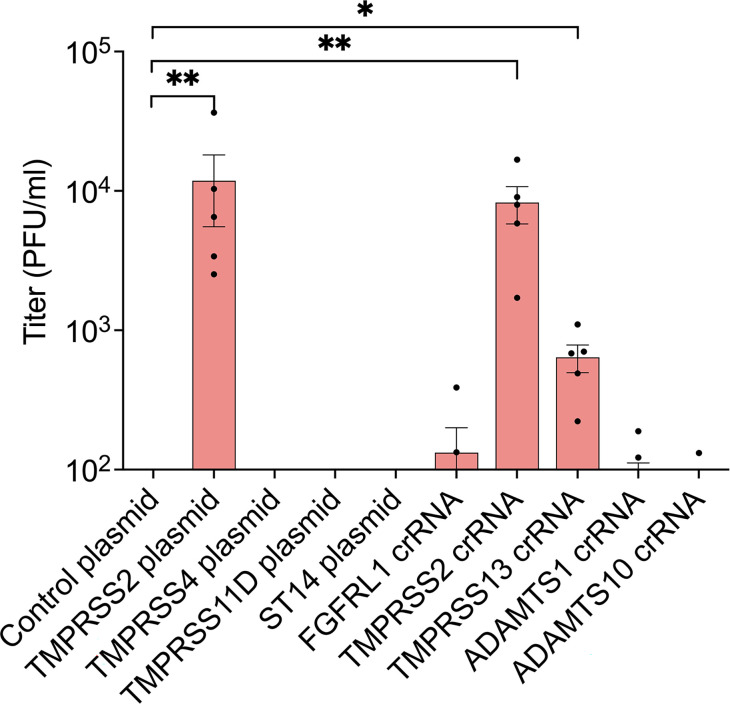
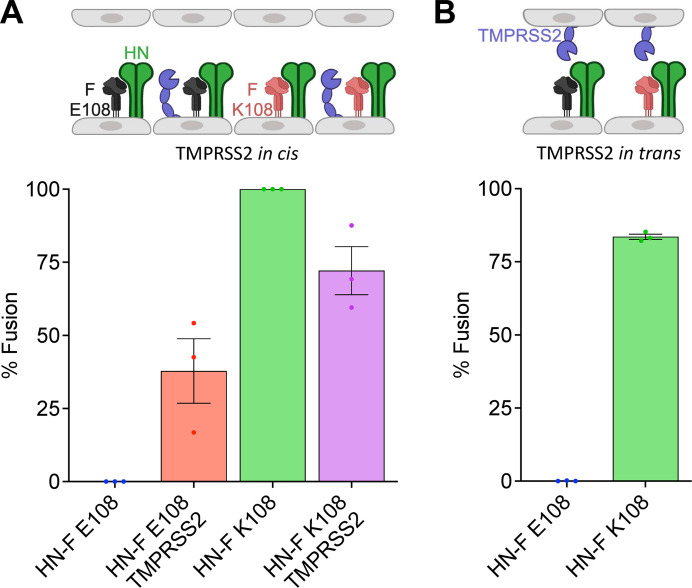
Human parainfluenza virus 3 (HPIV3) infection is driven by the coordinated action of viral surface glycoproteins hemagglutinin-neuraminidase (HN) and fusion protein (F). Receptor-engaged HN activates F to insert into the target cell membrane and drive virion-cell membrane fusion. For F to mediate entry, its precursor (F0) must first be cleaved by host proteases. F0 cleavage has been thought to be executed during viral glycoprotein transit through the trans-Golgi network by the ubiquitously expressed furin because F0 proteins of laboratory-adapted viruses contain a furin recognition dibasic cleavage motif RXKR around residue 108. Here, we show that the F proteins of field strains have a different cleavage motif from laboratory-adapted strains and are cleaved by unidentified proteases expressed in only a narrow subset of cell types. We demonstrate that extracellular serine protease inhibitors block HPIV3 F0 cleavage for field strains, suggesting F0 cleavage occurs at the cell surface facilitated by transmembrane proteases. Candidate proteases that may process HPIV3 F were identified by a genome-wide CRISPRa screen in HEK293/dCas9-VP64 + MPH cells. The lung-expressed extracellular serine proteases TMPRSS2 and TMPRSS13 are both sufficient to cleave HPIV3 F and enable infectious virus release by otherwise non-permissive cells. Our findings support an alternative mechanism of F activation , reliant on extracellular membrane-bound serine proteases expressed in a narrow subset of cells. The proportion of HPIV3 F proteins cleaved and infectious virus release is determined by host cell expression of requisite proteases, allowing just-in-time activation of F and positioning F cleavage as another key regulator of HPIV3 spread.
Enveloped viruses cause a wide range of diseases in humans. At the first step of infection, these viruses must fuse their envelope with a cell membrane to initiate infection. This fusion is mediated by viral proteins that require a critical activating cleavage event. It was previously thought that for parainfluenza virus 3, an important cause of respiratory disease and a representative of a group of important pathogens, this cleavage event was mediated by furin in the cell secretory pathways prior to formation of the virions. We show that this is only true for laboratory strain viruses, and that clinical viruses that infect humans utilize extracellular proteases that are only made by a small subset of cells. These results highlight the importance of studying authentic clinical viruses that infect human tissues for understanding natural infection.
人类副流感病毒 3(HPIV3)感染是由病毒表面糖蛋白血凝素神经氨酸酶(HN)和融合蛋白(F)的协调作用驱动的。受体结合的 HN 激活 F 插入靶细胞膜并驱动病毒包膜与细胞膜融合。为了介导进入,其前体(F0)必须首先被宿主蛋白酶切割。F0 切割被认为是通过普遍表达的弗林蛋白酶在病毒糖蛋白穿过高尔基体内运输过程中执行的,因为实验室适应的病毒的 F0 蛋白在残基 108 附近含有弗林识别的双碱性切割基序 RXKR。在这里,我们表明,现场分离株的 F 蛋白具有不同于实验室适应株的不同切割基序,并且由仅在一小部分细胞类型中表达的未鉴定蛋白酶切割。我们证明,细胞外丝氨酸蛋白酶抑制剂可阻断现场分离株的 HPIV3 F0 切割,这表明 F0 切割发生在细胞表面,由跨膜蛋白酶介导。通过 HEK293/dCas9-VP64+MPH 细胞的全基因组 CRISPRa 筛选鉴定了可能加工 HPIV3 F 的候选蛋白酶。肺表达的细胞外丝氨酸蛋白酶 TMPRSS2 和 TMPRSS13 都足以切割 HPIV3 F,并使原本不可感染的细胞释放感染性病毒。我们的发现支持 F 激活的替代机制,依赖于在一小部分细胞中表达的细胞外膜结合丝氨酸蛋白酶。F 蛋白的切割和感染性病毒的释放比例由宿主细胞所需蛋白酶的表达决定,这使得 F 的激活恰到好处,并将 F 切割定位为 HPIV3 传播的另一个关键调节剂。
包膜病毒可引起人类的广泛疾病。在感染的第一步,这些病毒必须使它们的包膜与细胞膜融合,以启动感染。这种融合由病毒蛋白介导,这些蛋白需要一个关键的激活切割事件。以前认为,对于副流感病毒 3,这是一种重要的呼吸道疾病病原体,也是一组重要病原体的代表,这种切割事件是在病毒形成之前在细胞分泌途径中由弗林蛋白酶介导的。我们表明,这仅适用于实验室株病毒,而感染人类的临床病毒则利用仅由一小部分细胞产生的细胞外蛋白酶。这些结果强调了研究感染人类组织的真实临床病毒对于理解自然感染的重要性。