Department of Orthopaedic Surgery, Stanford University, 450 Broadway Street, Redwood City, CA, 94025, USA.
Department of Orthopaedic Surgery, Stanford School of Medicine, Biomedical Innovations Building, 240 Pasteur Drive, Palo Alto, CA, 94304, USA.
J Transl Med. 2024 Nov 19;22(1):1041. doi: 10.1186/s12967-024-05866-5.
Diagnosing infected joint replacements relies heavily on assessing the neutrophil response to bacteria. Bacteria form biofilms on joint replacements. Biofilms are sessile bacterial communities encased in a protective extracellular matrix, making them notoriously difficult to culture, remarkably tolerant to antibiotics, and able to evade phagocytosis. Phagocytized bacteria dramatically alter cytokine production and compromise macrophage antigen presentation. We hypothesize that a subset of joint replacements have a dormant infection that suppresses the neutrophil response to bacteria but can be distinguished from uninfected joint replacements by the response of the mononuclear phagocyte system (MPS) within periarticular tissue, synovial fluid, and circulating plasma.
Single cell RNASeq transcriptomic and OLink proteomic profiling was performed on matched whole blood, synovial fluid, and periarticular tissue samples collected from 4 joint replacements with an active infection and 3 joint replacements without infection as well as 6 joint replacements with a prior infection deemed "infection-free" by the 2018 Musculoskeletal Infection Society criteria (follow-up of 26 ± 3 months).
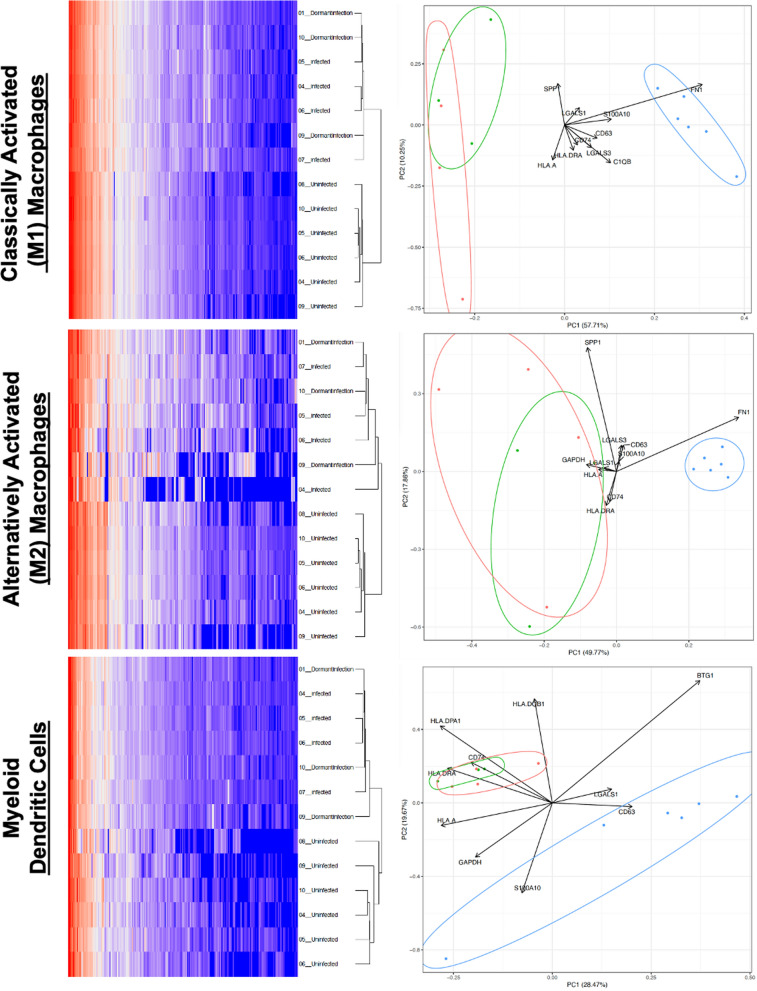
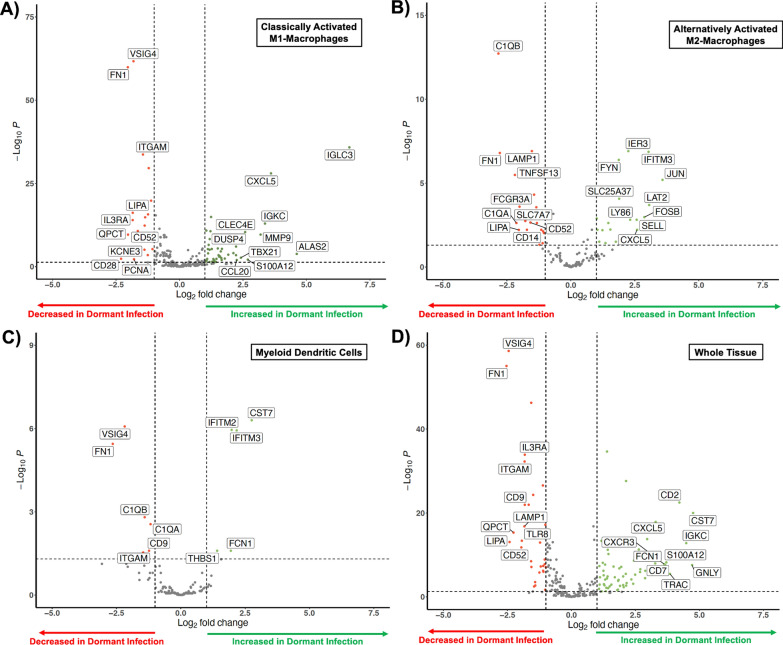
The MPS and neutrophil responses differ by infected state; the cellular distribution of the MPS response in the subset of joints with dormant infections resembled actively infected joints (p = 0.843, Chi-square test) but was significantly different from uninfected joints (p < 0.001, Chi-square test) despite the absence of systemic acute phase reactants and recruitment of neutrophils (p < 0.001, t-test). When compared to no infection, the cellular composition of dormant infection was distinct. There was reduction in classically activated M1 macrophages (p < 0.001, Fischer's test) and alternatively activated M2 macrophages coupled with an increase in classical monocytes (p < 0.001, Fischer's test), myeloid dendritic cells (p < 0.001, Fischer's test), regulatory T-cells (p < 0.001, Fischer's test), natural killer cells (p = 0.009, Fischer's test), and plasmacytoid dendritic cells (p = 0.005, Fischer's test). Hierarchical cluster analysis and single-cell gene expression revealed that classically M1 and alternatively M2 activated macrophages as well as myeloid dendritic cells can independently distinguish the dormant and uninfected patient populations suggesting that a process that modulates neutrophil recruitment (C1QA, C1QB, LY86, SELL, CXCL5, CCL20, CD14, ITGAM), macrophage polarization (FOSB, JUN), immune checkpoint regulation (IFITM2, IFITM3, CST7, THBS1), and T-cell response (VISIG4, CD28, FYN, LAT2, FCGR3A, CD52) was occurring during dormant infection. Gene set variation analysis suggested that activation of the TNF (FDR < 0.01) and IL17 (FDR < 0.01) pathways may distinguish dormant infections from the active and uninfected populations, while an inactivation of neutrophil extracellular traps (NETs) may be involved in the lack of a clinical response to a dormant infection using established diagnostic criteria. Synovial inflammatory proteomics show an increase in synovial CXCL5 associated with dormant infection (p = 0.011, t-test), suggesting the establishment of a chronic inflammatory state by the MPS during a dormant infection involved in neutrophil inhibition. Plasma inflammatory proteomics also support a chronic inflammatory state (EGF, GZMN, FGF2, PTN, MMP12) during dormant infection that involves a reduction in neutrophil recruitment (CXCL5, p = 0.006, t-test), antigen presentation (LAMP3, p = 0.047, t-test), and T-cell function (CD28, p = 0.045, t-test; CD70, p = 0.002, t-test) that are also seen during the development of bacterial tolerance.
All current diagnostic criteria assume each patient can mount the same neutrophil response to an implant-associated infection. However, the state of the MPS is of critical importance to accurate diagnosis of an implant-associated infection. A reduction in neutrophil recruitment and function mediated by the MPS may allow joint replacements with a dormant infection to be mischaracterized as uninfected, thus limiting the prognostic capabilities of all current diagnostic tests.
诊断感染性关节置换物严重依赖于评估对细菌的中性粒细胞反应。细菌在关节置换物上形成生物膜。生物膜是一种由保护性细胞外基质包裹的静止细菌群落,使其极难培养,对抗生素具有极强的耐受性,并能逃避吞噬作用。被吞噬的细菌会显著改变细胞因子的产生,并损害巨噬细胞的抗原呈递。我们假设一部分关节置换物存在休眠性感染,这种感染抑制了中性粒细胞对细菌的反应,但可以通过关节周围组织、滑液和循环血浆中单核吞噬细胞系统(MPS)的反应与未感染的关节置换物区分开来。
对来自 4 例活动性感染和 3 例无感染的关节置换物以及 6 例先前被认为符合 2018 年肌肉骨骼感染协会标准的“无感染”的关节置换物(随访 26±3 个月)的匹配全血、滑液和关节周围组织样本进行单细胞 RNA 测序转录组和 OLink 蛋白质组学分析。
MPS 和中性粒细胞反应因感染状态而异;在休眠性感染的关节亚群中,MPS 的细胞分布与活动性感染的关节相似(p=0.843,卡方检验),但与未感染的关节明显不同(p<0.001,卡方检验),尽管没有全身性急性期反应物和中性粒细胞的募集(p<0.001,t 检验)。与无感染相比,休眠性感染的细胞组成是不同的。经典激活的 M1 巨噬细胞(p<0.001,Fisher 检验)和替代激活的 M2 巨噬细胞减少,同时经典单核细胞增加(p<0.001,Fisher 检验),髓样树突状细胞(p<0.001,Fisher 检验),调节性 T 细胞(p<0.001,Fisher 检验),自然杀伤细胞(p=0.009,Fisher 检验)和浆细胞样树突状细胞(p=0.005,Fisher 检验)增加。层次聚类分析和单细胞基因表达显示,经典的 M1 和替代的 M2 激活的巨噬细胞以及髓样树突状细胞可以独立区分休眠和未感染的患者群体,这表明一种调节中性粒细胞募集的过程(C1QA、C1QB、LY86、SELL、CXCL5、CCL20、CD14、ITGAM)、巨噬细胞极化(FOSB、JUN)、免疫检查点调节(IFITM2、IFITM3、CST7、THBS1)和 T 细胞反应(VISIG4、CD28、FYN、LAT2、FCGR3A、CD52)在休眠性感染期间发生。基因集变异分析表明,TNF(FDR<0.01)和 IL17(FDR<0.01)途径的激活可能将休眠性感染与活动性和未感染人群区分开来,而中性粒细胞细胞外陷阱(NETs)的失活可能与使用现有诊断标准对休眠性感染缺乏临床反应有关。滑液炎症蛋白质组学显示,与休眠感染相关的滑液 CXCL5 增加(p=0.011,t 检验),这表明在休眠感染期间,MPS 建立了一种涉及中性粒细胞抑制的慢性炎症状态。血浆炎症蛋白质组学也支持休眠感染时的慢性炎症状态(EGF、GZMN、FGF2、PTN、MMP12),这涉及到中性粒细胞募集的减少(CXCL5,p=0.006,t 检验)、抗原呈递(LAMP3,p=0.047,t 检验)和 T 细胞功能(CD28,p=0.045,t 检验;CD70,p=0.002,t 检验),这些也见于细菌耐受的发展。
所有现有的诊断标准都假设每个患者都能对植入物相关感染产生相同的中性粒细胞反应。然而,MPS 的状态对植入物相关感染的准确诊断至关重要。由 MPS 介导的中性粒细胞募集和功能的减少可能会导致带有休眠性感染的关节置换物被误诊为未感染,从而限制了所有现有诊断测试的预后能力。