Zhang Shuai, Wang Ruixue, Luo Kun, Gu Shipeng, Liu Xinyu, Wang Junhan, Zhang Ludan, Sun Lin
Key Laboratory of Cell Proliferation and Regulation Biology of Ministry of Education, College of Life Sciences, Beijing Normal University, Beijing, China.
Beijing Key Laboratory of Gene Resource and Molecular Development, College of Life Sciences, Beijing Normal University, Beijing, China.
Elife. 2025 Jan 24;13:RP100144. doi: 10.7554/eLife.100144.
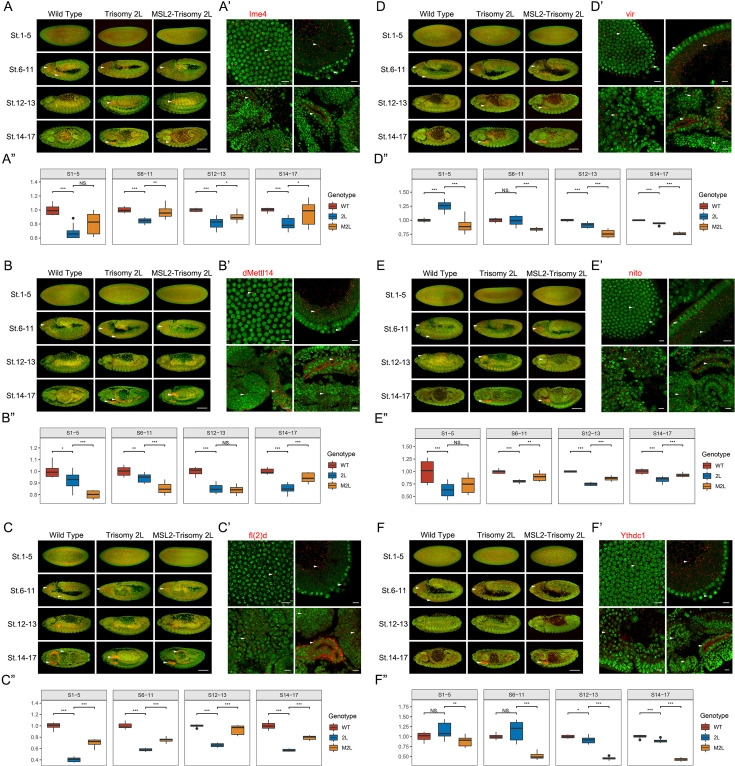
-methyladenosine (mA) in eukaryotic RNA is an epigenetic modification that is critical for RNA metabolism, gene expression regulation, and the development of organisms. Aberrant expression of mA components appears in a variety of human diseases. RNA mA modification in has proven to be involved in sex determination regulated by and may affect X chromosome expression through the MSL complex. The dosage-related effects under the condition of genomic imbalance (i.e. aneuploidy) are related to various epigenetic regulatory mechanisms. Here, we investigated the roles of RNA mA modification in unbalanced genomes using aneuploid . The results showed that the expression of mA components changed significantly under genomic imbalance, and affected the abundance and genome-wide distribution of mA, which may be related to the developmental abnormalities of aneuploids. The relationships between methylation status and classical dosage effect, dosage compensation, and inverse dosage effect were also studied. In addition, we demonstrated that RNA mA methylation may affect dosage-dependent gene regulation through dosage-sensitive modifiers, alternative splicing, the MSL complex, and other processes. More interestingly, there seems to be a close relationship between MSL complex and RNA mA modification. It is found that ectopically overexpressed MSL complex, especially the levels of H4K16Ac through MOF, could influence the expression levels of mA modification and genomic imbalance may be involved in this interaction. We found that mA could affect the levels of H4K16Ac through MOF, a component of the MSL complex, and that genomic imbalance may be involved in this interaction. Altogether, our work reveals the dynamic and regulatory role of RNA mA modification in unbalanced genomes, and may shed new light on the mechanisms of aneuploidy-related developmental abnormalities and diseases.
真核生物RNA中的N6-甲基腺苷(mA)是一种表观遗传修饰,对RNA代谢、基因表达调控和生物体发育至关重要。mA组分的异常表达出现在多种人类疾病中。已证明RNA mA修饰参与由[未提及具体因素]调控的性别决定,并可能通过MSL复合体影响X染色体表达。基因组失衡(即非整倍体)条件下的剂量相关效应与各种表观遗传调控机制有关。在这里,我们使用非整倍体研究了RNA mA修饰在不平衡基因组中的作用。结果表明,在基因组失衡情况下,mA组分的表达发生了显著变化,并影响了mA的丰度和全基因组分布,这可能与非整倍体的发育异常有关。还研究了甲基化状态与经典剂量效应、剂量补偿和反剂量效应之间的关系。此外,我们证明RNA mA甲基化可能通过剂量敏感修饰因子、可变剪接、MSL复合体和其他过程影响剂量依赖性基因调控。更有趣的是,MSL复合体与RNA mA修饰之间似乎存在密切关系。发现异位过表达的MSL复合体,特别是通过MOF导致的H4K16Ac水平,可影响mA修饰的表达水平,基因组失衡可能参与了这种相互作用。我们发现mA可通过MSL复合体的组分MOF影响H4K16Ac水平,基因组失衡可能参与了这种相互作用。总之,我们的工作揭示了RNA mA修饰在不平衡基因组中的动态和调控作用,并可能为非整倍体相关发育异常和疾病的机制提供新的线索。