Graceffa P, Adam L P, Lehman W
Department of Muscle Research, Boston Biomedical Research Institute, MA 02114.
Biochem J. 1993 Aug 15;294 ( Pt 1)(Pt 1):63-7. doi: 10.1042/bj2940063.
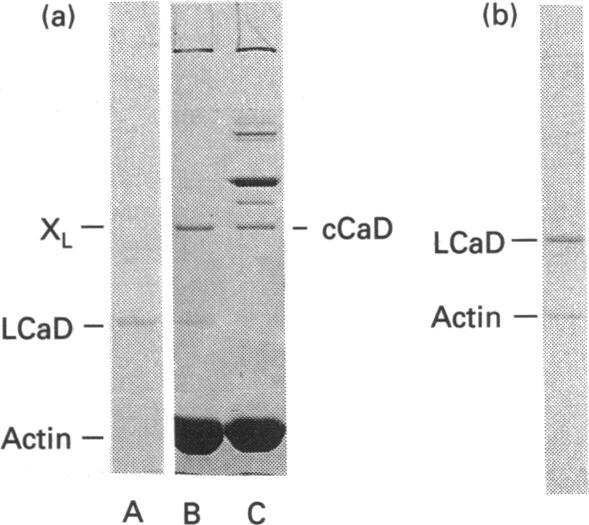
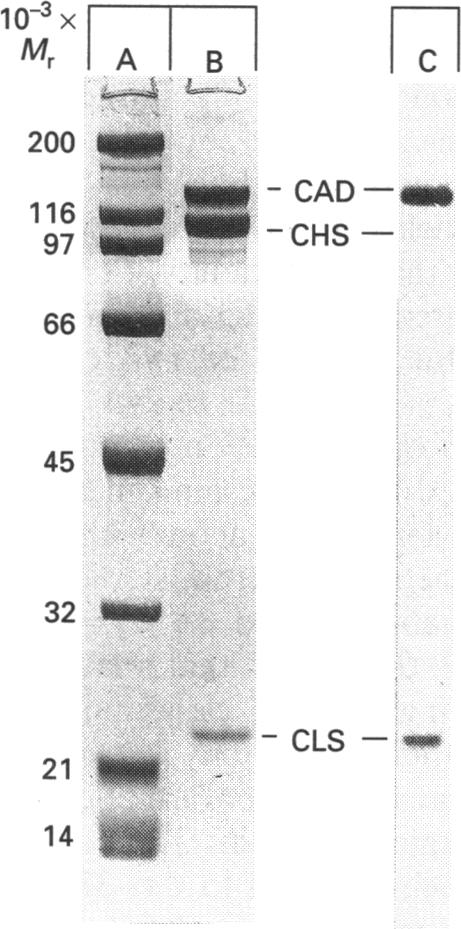
It was reported that chicken gizzard smooth-muscle caldesmon Cys-580 can be disulphide-cross-linked to the C-terminal pen-ultimate residue (Cys-374) of actin, indicating that these residues are close in the protein complex [Graceffa, P. and Jancso, A. (1991) J. Biol. Chem. 266, 20305-20310]. Since the possibility that the cross-link involves a cysteine residue other than actin Cys-374 was not absolutely excluded, more direct evidence was sought for the identify of the cysteine residues involved in the cross-link. We show here that caldesmon could not be disulphide-cross-linked to actin which had Cys-374 removed by carboxypeptidase A digestion, providing direct support for the participation of actin Cys-374 in the cross-link to caldesmon. In order to assign the caldesmon cysteine residue involved in the cross-link, use was made of caldesmon from porcine stomach muscle, which is shown to contain one cysteine residue close to, or at, position 580, in contrast with chicken gizzard caldesmon, which has an additional cysteine residue at position 153. The porcine stomach caldesmon also formed a disulphide-cross-link to actin, further supporting the original conclusion that Cys-580 of the chicken gizzard caldesmon had been cross-linked to actin. Disulphide-cross-linking with similar yield was also observed in native chicken gizzard muscle thin filaments, indicating that the interaction between actin and the C-terminal domain of caldesmon is the same in native and reconstituted thin filaments. The much smaller non-muscle isoform of caldesmon, from rabbit liver, could be similarly cross-linked to actin, consistent with the sequence similarity between the C-terminal domain of muscle and non-muscle caldesmon. The ability to cross-link caldesmon Cys-580 to actin Cys-374 suggests the possibility that the Cys-580 region of caldesmon and the C-terminus of actin form part of the actin-caldesmon binding interface.
据报道,鸡胗平滑肌钙调蛋白的半胱氨酸-580可与肌动蛋白的C末端倒数第二个残基(半胱氨酸-374)形成二硫键交联,这表明在蛋白质复合物中这些残基距离较近[格雷斯法,P.和扬乔,A.(1991年)《生物化学杂志》266,20305 - 20310]。由于不能完全排除交联涉及肌动蛋白半胱氨酸-374以外的半胱氨酸残基的可能性,因此需要寻找更直接的证据来确定参与交联的半胱氨酸残基。我们在此表明,经羧肽酶A消化去除半胱氨酸-374的肌动蛋白不能与钙调蛋白形成二硫键交联,这为肌动蛋白半胱氨酸-374参与与钙调蛋白的交联提供了直接支持。为了确定参与交联的钙调蛋白半胱氨酸残基,使用了来自猪胃肌的钙调蛋白,已表明其在靠近或处于位置(580)处含有一个半胱氨酸残基,与之形成对比的是鸡胗钙调蛋白在位置(153)处还有一个额外的半胱氨酸残基。猪胃钙调蛋白也与肌动蛋白形成了二硫键交联,进一步支持了最初的结论,即鸡胗钙调蛋白的半胱氨酸-580已与肌动蛋白交联。在天然鸡胗肌细肌丝中也观察到了产率相似的二硫键交联,这表明在天然和重组细肌丝中肌动蛋白与钙调蛋白C末端结构域之间的相互作用是相同的。来自兔肝的小得多的非肌肉型钙调蛋白异构体也能类似地与肌动蛋白交联,这与肌肉型和非肌肉型钙调蛋白C末端结构域之间的序列相似性一致。将钙调蛋白半胱氨酸-580与肌动蛋白半胱氨酸-374交联的能力表明,钙调蛋白的半胱氨酸-580区域和肌动蛋白的C末端可能构成肌动蛋白 - 钙调蛋白结合界面的一部分。