Rosenberg A R, Delamarre L, Pique C, Le Blanc I, Griffith G, Dokhélar M C
Institut National de la Santé et de la Recherche Medicale U332, Institut Cochin de Génétique Moléculaire, 75014 Paris, France.
J Cell Biol. 1999 Apr 5;145(1):57-68. doi: 10.1083/jcb.145.1.57.
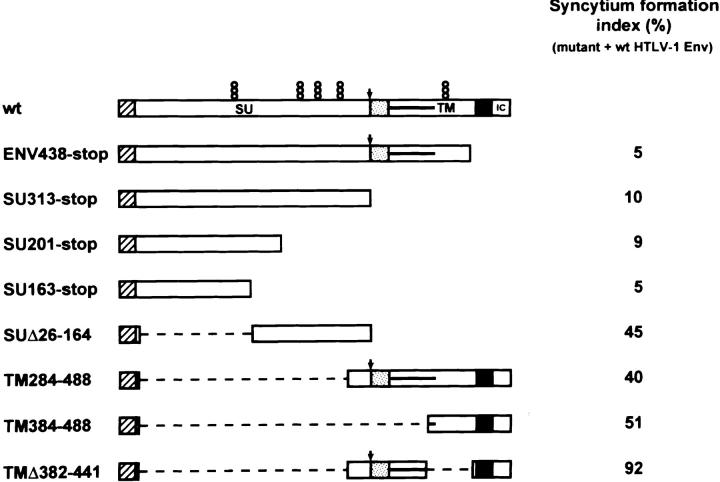
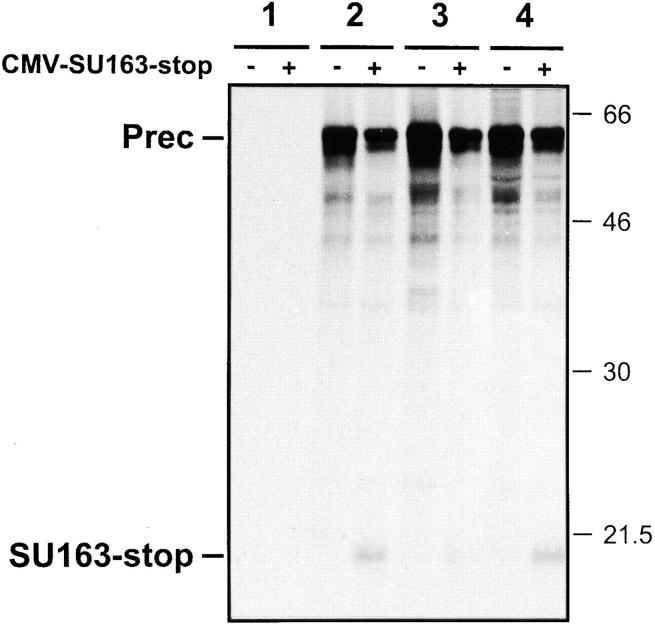
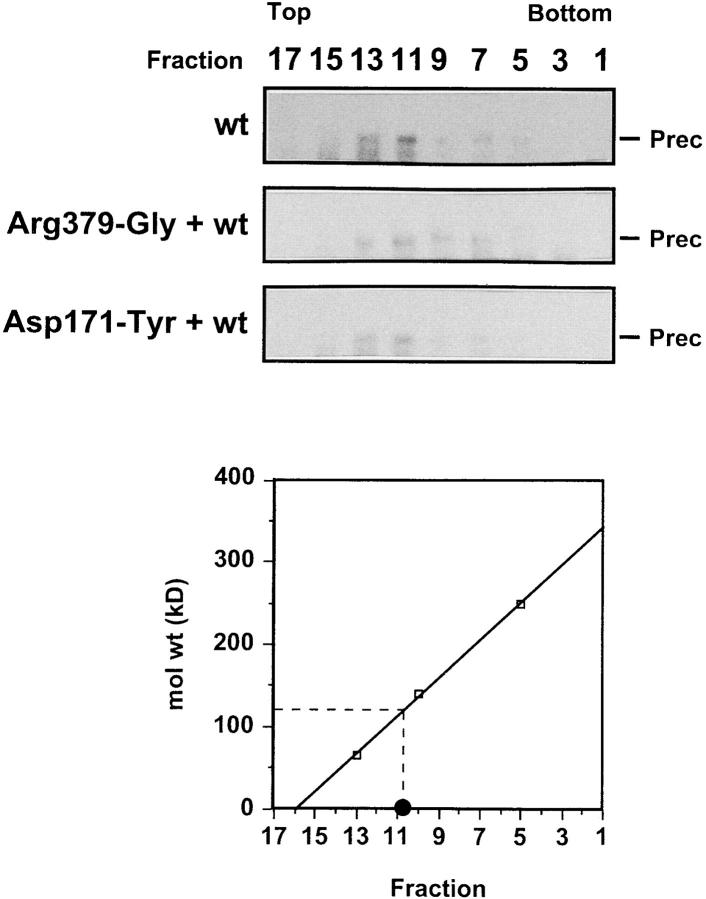
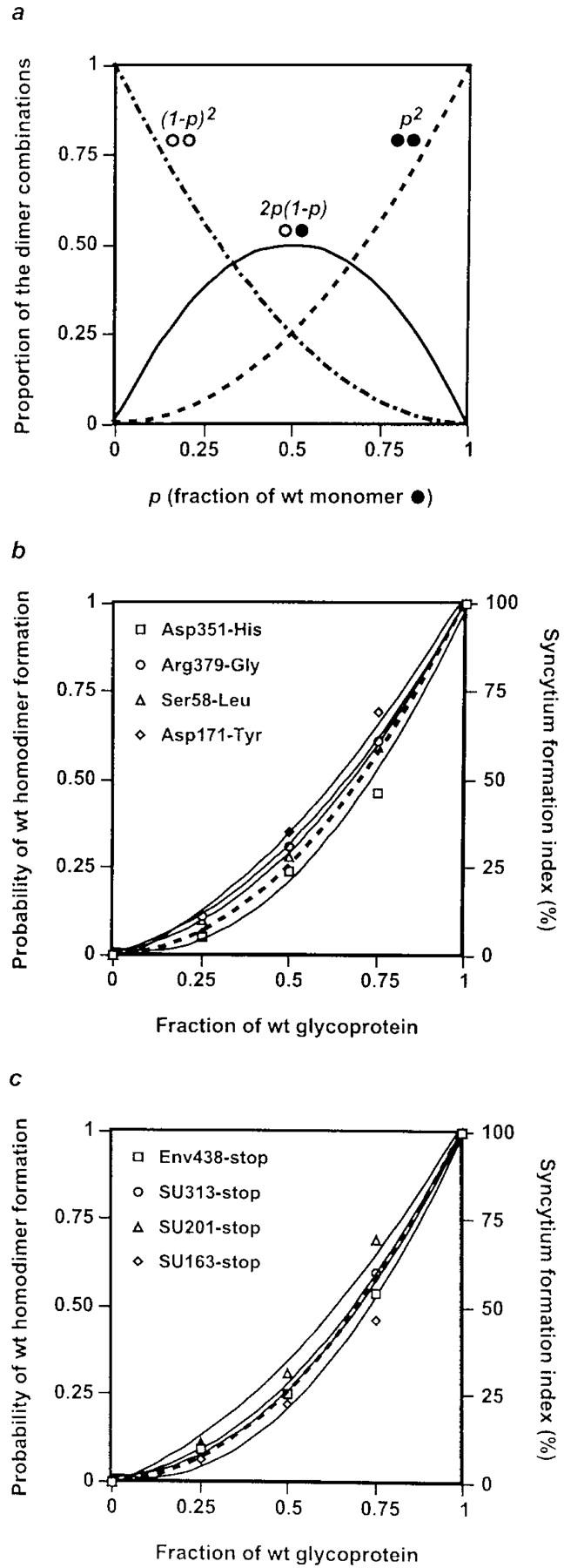
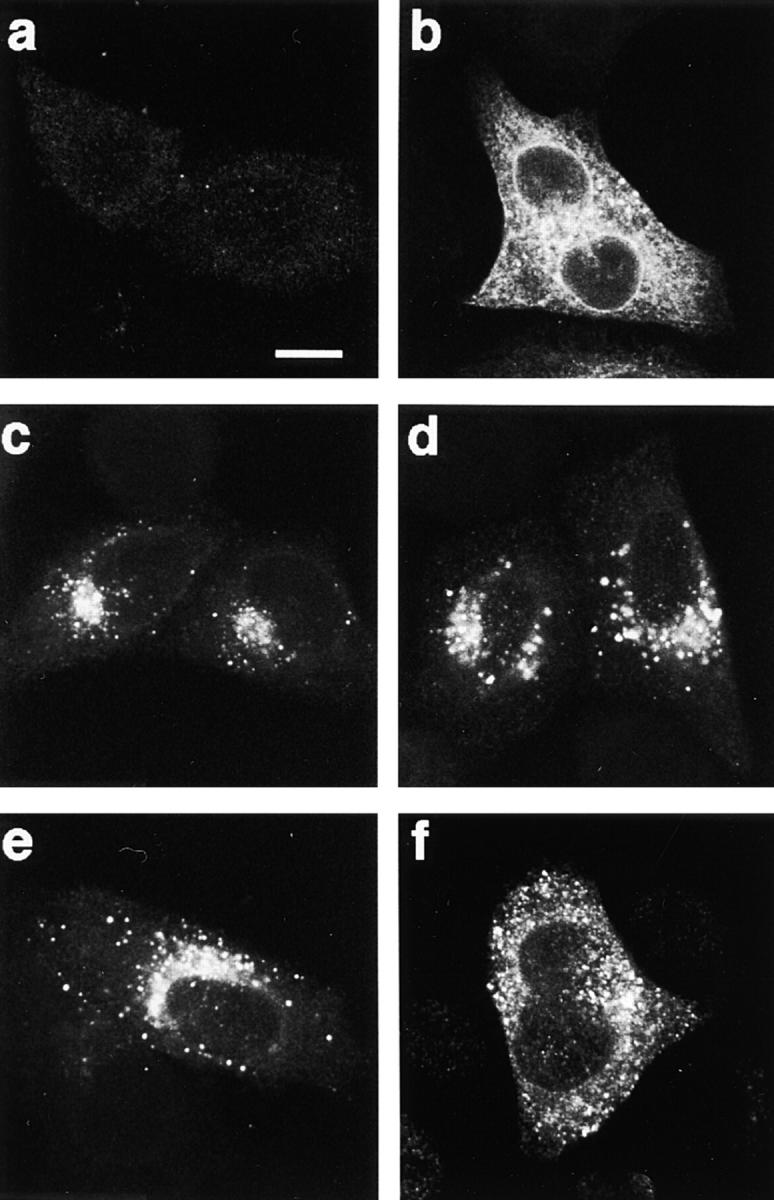
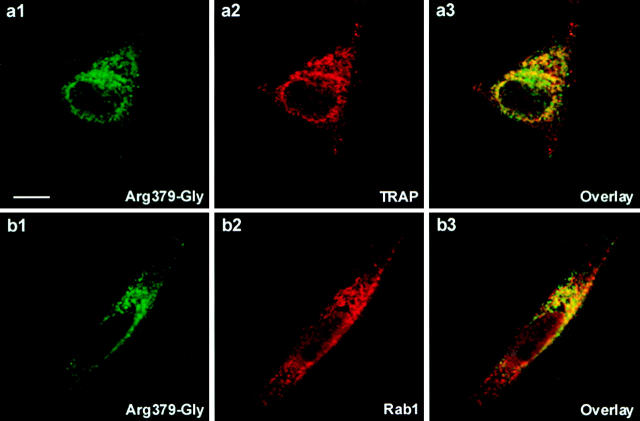
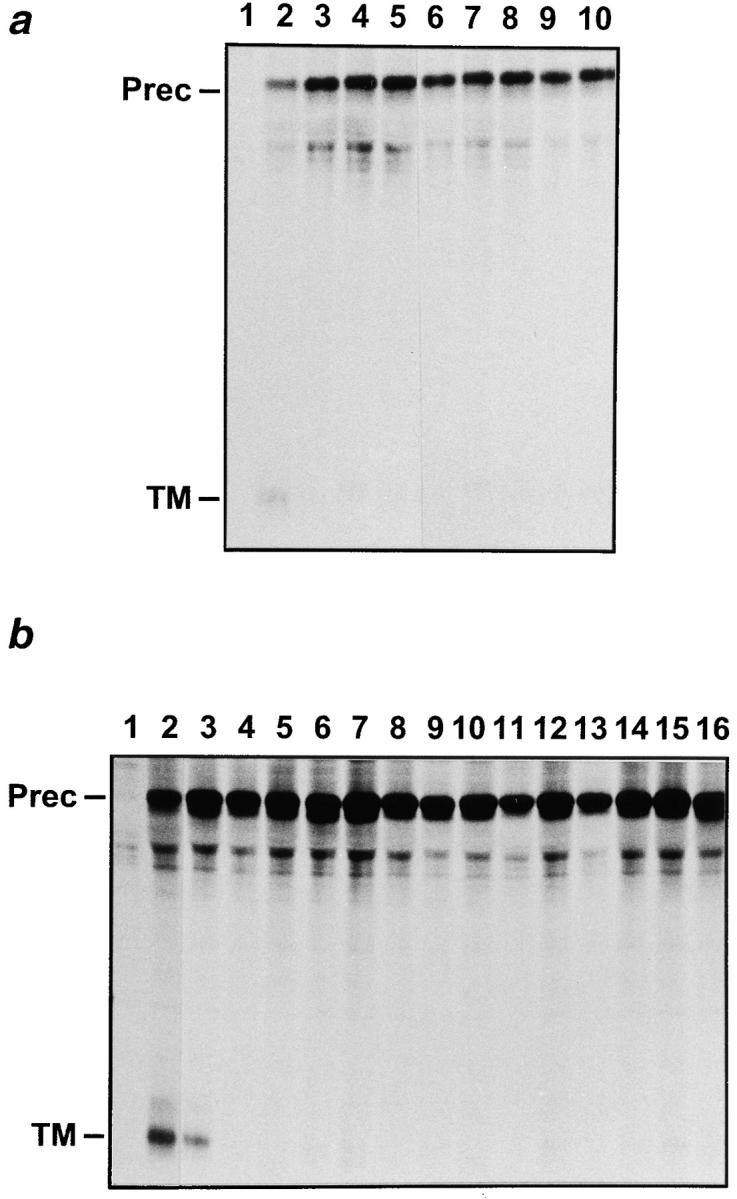
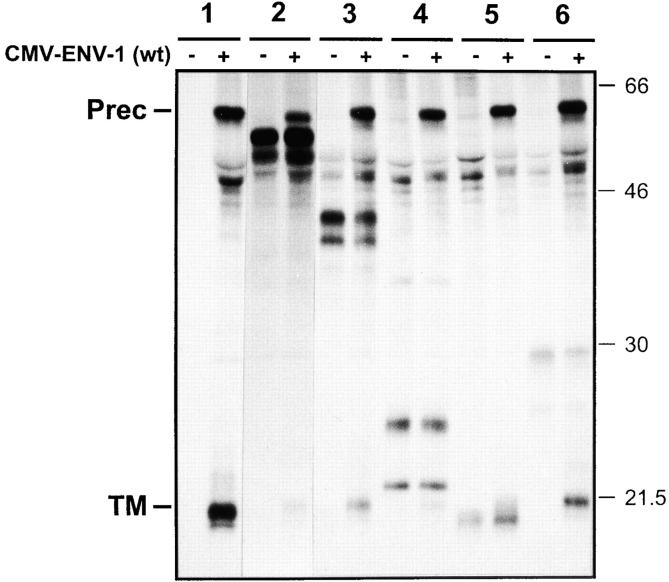
As for most integral membrane proteins, the intracellular transport of retroviral envelope glycoproteins depends on proper folding and oligomeric assembly in the ER. In this study, we considered the hypothesis that a panel of 22 transport-defective mutants of the human T cell leukemia virus type 1 envelope glycoprotein might be defective in ER assembly. Upon cell cotransfection with wild-type envelope, however, the vast majority of these transport-defective mutants (21 of 22) exerted a specific trans-dominant negative effect. This effect was due to random dimerization of the mutated and wild-type glycoproteins that prevented the intracellular transport of the latter. This unexpected result suggests that association of glycoprotein monomers precedes the completion of folding. The only mutation that impaired this early assembly was located at the NH2 terminus of the protein. COOH-terminally truncated, soluble forms of the glycoprotein were also trans-dominant negative provided that their NH2 terminus was intact. The leucine zipper-like domain, although involved in oligomerization of the envelope glycoproteins at the cell surface, did not contribute to their intracellular assembly. We propose that, at a step subsequent to translation, but preceding complete folding of the monomers, glycoproteins assemble via their NH2-terminal domains, which, in turn, permits their cooperative folding.
对于大多数整合膜蛋白而言,逆转录病毒包膜糖蛋白的细胞内运输依赖于其在内质网中的正确折叠和寡聚体组装。在本研究中,我们考虑了这样一种假说,即一组22个1型人类T细胞白血病病毒包膜糖蛋白的运输缺陷型突变体可能在内质网组装方面存在缺陷。然而,当与野生型包膜进行细胞共转染时,这些运输缺陷型突变体中的绝大多数(22个中的21个)发挥了特异性的反式显性负效应。这种效应是由于突变型和野生型糖蛋白的随机二聚化阻止了后者的细胞内运输。这一意外结果表明糖蛋白单体的缔合先于折叠的完成。唯一损害这种早期组装的突变位于该蛋白的NH2末端。只要其NH2末端完整,糖蛋白的COOH末端截短的可溶性形式也具有反式显性负效应。亮氨酸拉链样结构域虽然参与包膜糖蛋白在细胞表面的寡聚化,但对其细胞内组装没有贡献。我们提出,在翻译后的一个步骤中,但在单体完全折叠之前,糖蛋白通过其NH2末端结构域进行组装,这反过来又允许它们协同折叠。