Yao Jun, Zhong Jin, Lambowitz Alan M
Institute for Cellular and Molecular Biology, Department of Chemistry and Biochemistry, and Section of Molecular Genetics and Microbiology, School of Biological Sciences, University of Texas at Austin Austin, TX 78712, USA.
Nucleic Acids Res. 2005 Jun 9;33(10):3351-62. doi: 10.1093/nar/gki649. Print 2005.
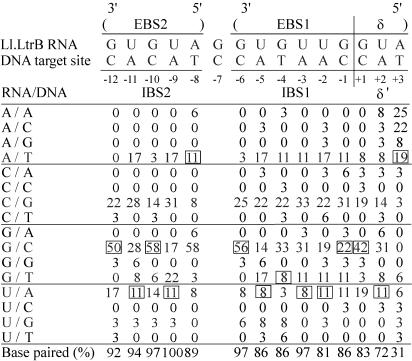
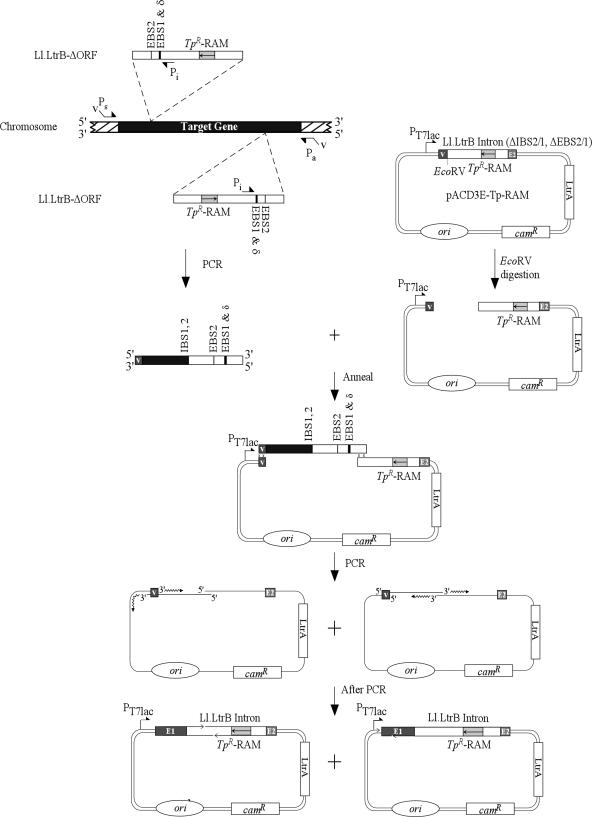
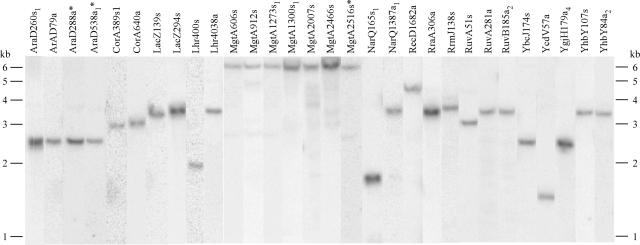
The Lactococcus lactis Ll.LtrB group II intron retrohomes by reverse-splicing into one strand of a double-stranded DNA target site, while the intron-encoded protein cleaves the opposite strand and uses it to prime reverse transcription of the inserted intron RNA. The protein and intron RNA function in a ribonucleoprotein particle, with much of the DNA target sequence recognized by base-pairing of the intron RNA. Consequently, group II introns can be reprogrammed to insert into specific or random DNA sites by substituting specific or random nucleotide residues in the intron RNA. Here, we show that an Escherichia coli gene disruption library obtained using such randomly inserting Ll.LtrB introns contains most viable E.coli gene disruptions. Further, each inserted intron is targeted to a specific site by its unique base-pairing regions, and in most cases, could be recovered by PCR and used unmodified to obtain the desired single disruptant. Additionally, we identified a subset of introns that insert at sites lacking T+5, a nucleotide residue critical for second-strand cleavage. All such introns tested individually gave the desired specific disruption, some by switching to an alternate retrohoming mechanism targeting single-stranded DNA and using a nascent lagging DNA strand to prime reverse transcription.
乳酸乳球菌Ll.LtrB II组内含子通过反向剪接归巢到双链DNA靶位点的一条链中,而内含子编码的蛋白质切割另一条链并利用它引发插入的内含子RNA的逆转录。该蛋白质和内含子RNA在核糖核蛋白颗粒中发挥作用,大部分DNA靶序列通过内含子RNA的碱基配对识别。因此,通过替换内含子RNA中的特定或随机核苷酸残基,II组内含子可以被重新编程以插入特定或随机的DNA位点。在这里,我们表明,使用这种随机插入的Ll.LtrB内含子获得的大肠杆菌基因破坏文库包含了大多数可行的大肠杆菌基因破坏。此外,每个插入的内含子通过其独特的碱基配对区域靶向特定位点,并且在大多数情况下,可以通过PCR回收并未经修饰地用于获得所需的单破坏体。此外,我们鉴定了一组内含子,它们插入缺乏T+5的位点,T+5是第二链切割的关键核苷酸残基。单独测试的所有此类内含子都产生了所需的特异性破坏,有些是通过切换到靶向单链DNA的另一种归巢机制并使用新生的滞后DNA链引发逆转录。