Schmid Eva M, Ford Marijn G J, Burtey Anne, Praefcke Gerrit J K, Peak-Chew Sew-Yeu, Mills Ian G, Benmerah Alexandre, McMahon Harvey T
Medical Research Council Laboratory of Molecular Biology, Cambridge, United Kingdom.
PLoS Biol. 2006 Sep;4(9):e262. doi: 10.1371/journal.pbio.0040262.
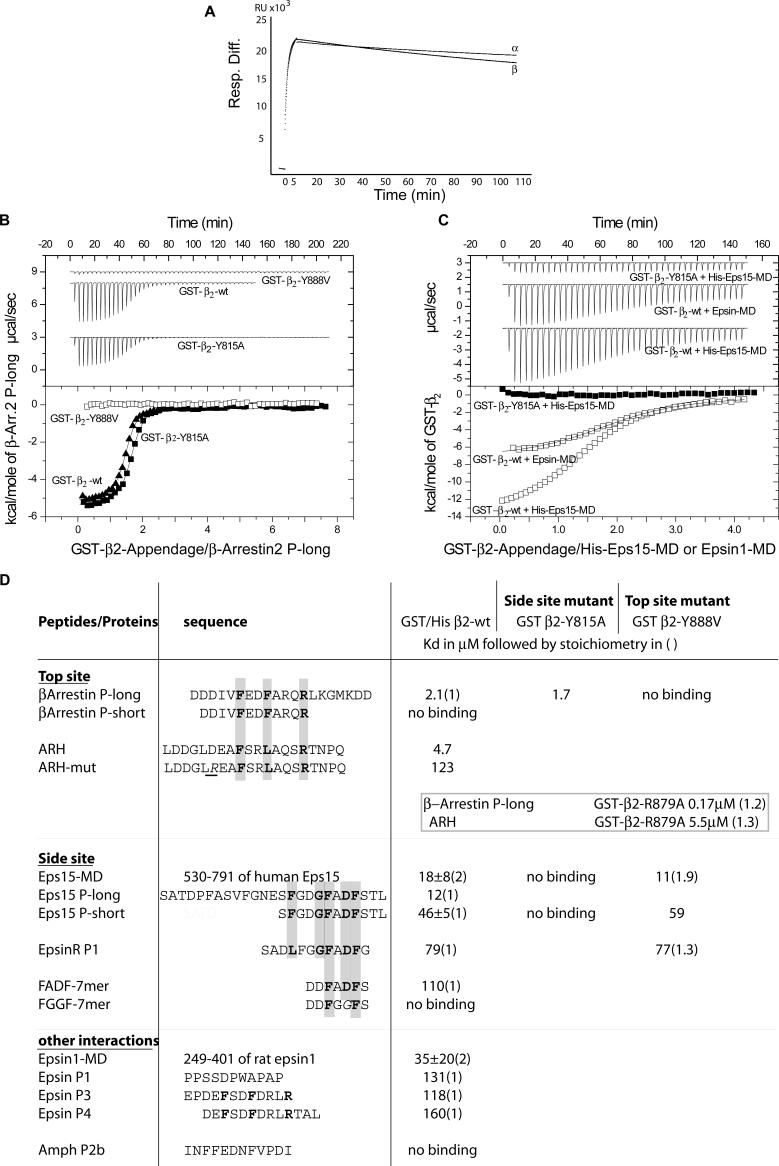
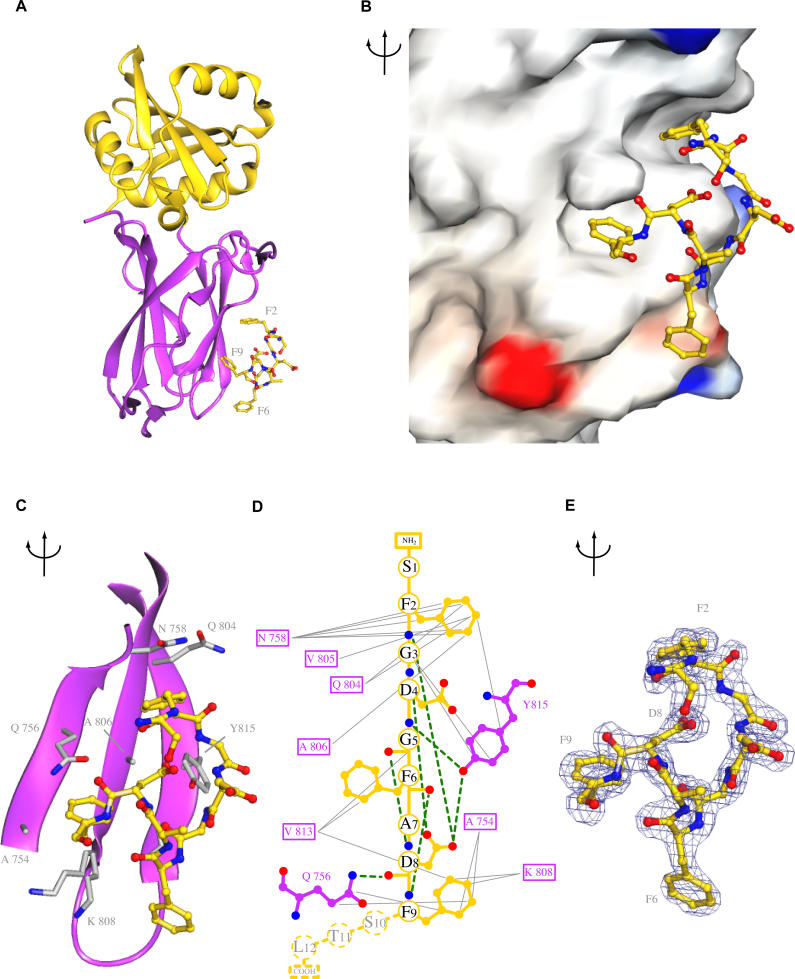
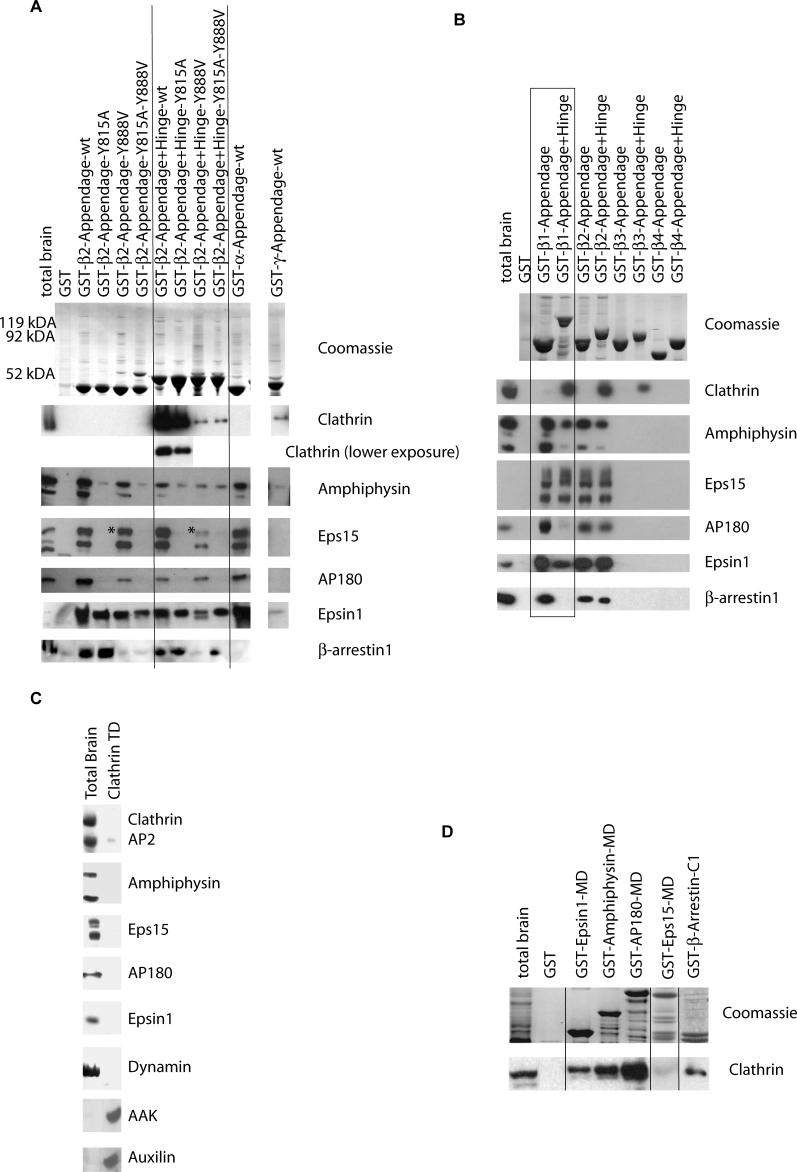
Adaptor protein complex 2 alpha and beta-appendage domains act as hubs for the assembly of accessory protein networks involved in clathrin-coated vesicle formation. We identify a large repertoire of beta-appendage interactors by mass spectrometry. These interact with two distinct ligand interaction sites on the beta-appendage (the "top" and "side" sites) that bind motifs distinct from those previously identified on the alpha-appendage. We solved the structure of the beta-appendage with a peptide from the accessory protein Eps15 bound to the side site and with a peptide from the accessory cargo adaptor beta-arrestin bound to the top site. We show that accessory proteins can bind simultaneously to multiple appendages, allowing these to cooperate in enhancing ligand avidities that appear to be irreversible in vitro. We now propose that clathrin, which interacts with the beta-appendage, achieves ligand displacement in vivo by self-polymerisation as the coated pit matures. This changes the interaction environment from liquid-phase, affinity-driven interactions, to interactions driven by solid-phase stability ("matricity"). Accessory proteins that interact solely with the appendages are thereby displaced to areas of the coated pit where clathrin has not yet polymerised. However, proteins such as beta-arrestin (non-visual arrestin) and autosomal recessive hypercholesterolemia protein, which have direct clathrin interactions, will remain in the coated pits with their interacting receptors.
衔接蛋白复合体2的α和β附器结构域作为参与网格蛋白包被小泡形成的辅助蛋白网络组装的枢纽。我们通过质谱鉴定了大量β附器相互作用蛋白。这些蛋白与β附器上两个不同的配体相互作用位点(“顶部”和“侧面”位点)相互作用,这些位点结合的基序与先前在α附器上鉴定的基序不同。我们解析了β附器与结合在侧面位点的辅助蛋白Eps15的肽段以及结合在顶部位点的辅助货物衔接蛋白β-抑制蛋白的肽段形成的复合物的结构。我们发现辅助蛋白可以同时结合多个附器,使它们协同增强配体亲和力,这种亲和力在体外似乎是不可逆的。我们现在提出,与β附器相互作用的网格蛋白在体内通过随着包被小窝成熟而进行的自我聚合来实现配体置换。这将相互作用环境从液相的、亲和力驱动的相互作用转变为由固相稳定性(“基质性”)驱动的相互作用。仅与附器相互作用的辅助蛋白因此被置换到包被小窝中网格蛋白尚未聚合的区域。然而,诸如β-抑制蛋白(非视觉抑制蛋白)和常染色体隐性高胆固醇血症蛋白等与网格蛋白直接相互作用的蛋白,将与其相互作用的受体一起留在包被小窝中。