Lowe Christopher J, Terasaki Mark, Wu Michael, Freeman Robert M, Runft Linda, Kwan Kristen, Haigo Saori, Aronowicz Jochanan, Lander Eric, Gruber Chris, Smith Mark, Kirschner Marc, Gerhart John
Department of Organismal Biology and Anatomy, University of Chicago, Chicago, Illinois, United States of America.
PLoS Biol. 2006 Sep;4(9):e291. doi: 10.1371/journal.pbio.0040291.
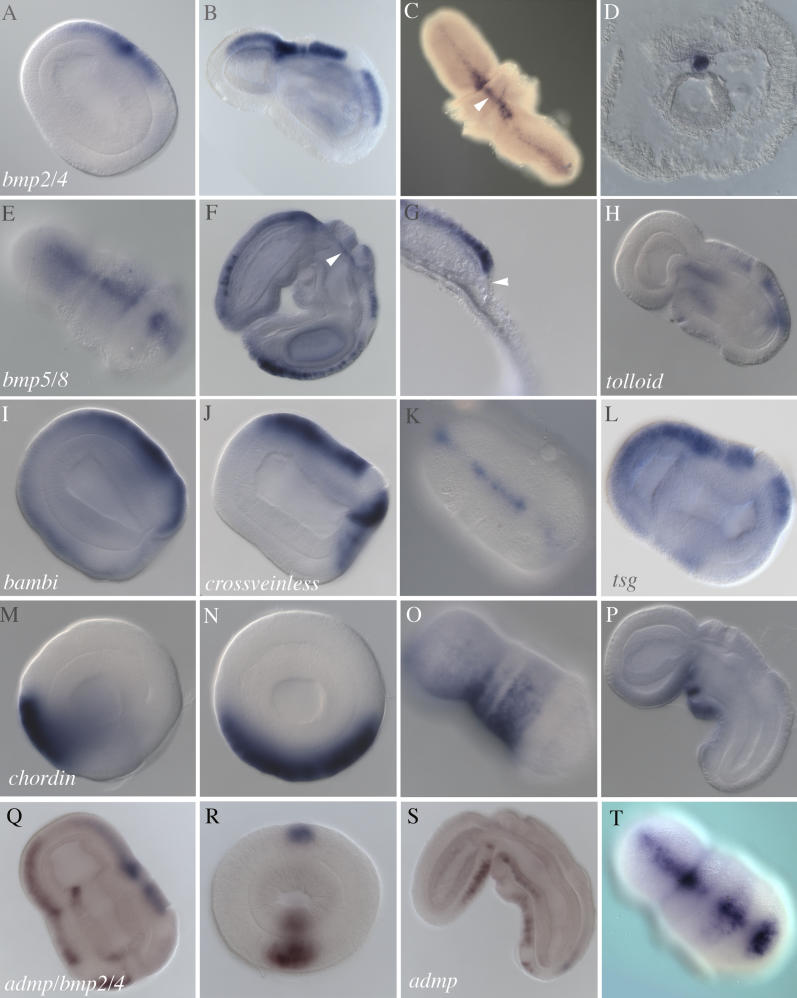
We have compared the dorsoventral development of hemichordates and chordates to deduce the organization of their common ancestor, and hence to identify the evolutionary modifications of the chordate body axis after the lineages split. In the hemichordate embryo, genes encoding bone morphogenetic proteins (Bmp) 2/4 and 5/8, as well as several genes for modulators of Bmp activity, are expressed in a thin stripe of ectoderm on one midline, historically called "dorsal." On the opposite midline, the genes encoding Chordin and Anti-dorsalizing morphogenetic protein (Admp) are expressed. Thus, we find a Bmp-Chordin developmental axis preceding and underlying the anatomical dorsoventral axis of hemichordates, adding to the evidence from Drosophila and chordates that this axis may be at least as ancient as the first bilateral animals. Numerous genes encoding transcription factors and signaling ligands are expressed in the three germ layers of hemichordate embryos in distinct dorsoventral domains, such as pox neuro, pituitary homeobox, distalless, and tbx2/3 on the Bmp side and netrin, mnx, mox, and single-minded on the Chordin-Admp side. When we expose the embryo to excess Bmp protein, or when we deplete endogenous Bmp by small interfering RNA injections, these expression domains expand or contract, reflecting their activation or repression by Bmp, and the embryos develop as dorsalized or ventralized limit forms. Dorsoventral patterning is independent of anterior/posterior patterning, as in Drosophila but not chordates. Unlike both chordates and Drosophila, neural gene expression in hemichordates is not repressed by high Bmp levels, consistent with their development of a diffuse rather than centralized nervous system. We suggest that the common ancestor of hemichordates and chordates did not use its Bmp-Chordin axis to segregate epidermal and neural ectoderm but to pattern many other dorsoventral aspects of the germ layers, including neural cell fates within a diffuse nervous system. Accordingly, centralization was added in the chordate line by neural-epidermal segregation, mediated by the pre-existing Bmp-Chordin axis. Finally, since hemichordates develop the mouth on the non-Bmp side, like arthropods but opposite to chordates, the mouth and Bmp-Chordin axis may have rearranged in the chordate line, one relative to the other.
我们比较了半索动物和脊索动物的背腹发育过程,以推断它们共同祖先的组织结构,从而确定在谱系分化后脊索动物体轴的进化变化。在半索动物胚胎中,编码骨形态发生蛋白(Bmp)2/4和5/8的基因,以及几个Bmp活性调节因子的基因,在一条中线的外胚层细条带中表达,这条中线在历史上被称为“背侧”。在相对的中线上,编码脊索蛋白和抗背化形态发生蛋白(Admp)的基因表达。因此,我们发现半索动物解剖学上的背腹轴之前存在一个Bmp - 脊索蛋白发育轴,这进一步证明了来自果蝇和脊索动物的证据,即这个轴可能至少与第一批两侧对称动物一样古老。许多编码转录因子和信号配体的基因在半索动物胚胎的三个胚层中,在不同的背腹区域表达,例如在Bmp一侧的痘神经、垂体同源框、远端缺失和tbx2/3,以及在脊索蛋白 - Admp一侧的网蛋白、mnx、mox和单-minded。当我们让胚胎暴露于过量的Bmp蛋白中,或者当我们通过注射小干扰RNA耗尽内源性Bmp时,这些表达区域会扩大或收缩,反映出它们被Bmp激活或抑制,并且胚胎发育为背化或腹化的极限形式。背腹模式形成独立于前后模式形成,这与果蝇相同,但与脊索动物不同。与脊索动物和果蝇都不同的是,半索动物中的神经基因表达不会被高Bmp水平抑制,这与它们发育出分散而非集中的神经系统一致。我们认为,半索动物和脊索动物的共同祖先并没有利用其Bmp - 脊索蛋白轴来分离表皮和神经外胚层,而是用于构建胚层的许多其他背腹方面,包括分散神经系统内的神经细胞命运。因此,在脊索动物谱系中,通过由预先存在的Bmp - 脊索蛋白轴介导的神经 - 表皮分离增加了集中化。最后,由于半索动物在非Bmp侧发育口,这与节肢动物类似,但与脊索动物相反,口和Bmp - 脊索蛋白轴在脊索动物谱系中可能相对于彼此发生了重排。