Walter Brian E, Perry Kimberly J, Fukui Lisa, Malloch Erica L, Wever Jason, Henry Jonathan J
Biology Department, Illinois Wesleyan University, Bloomington, IL, USA.
Mol Vis. 2008 May 19;14:906-21.
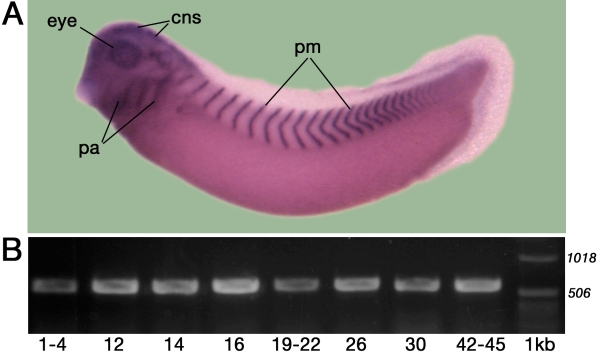
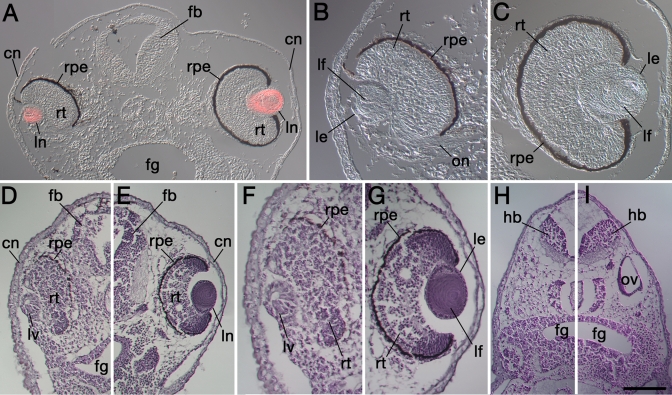
Psf2 (partner of Sld5 2) represents a member of the GINS (go, ichi, ni, san) heterotetramer [1] and functions in DNA replication as a "sliding clamp." Previous in situ hybridization analyses revealed that Psf2 is expressed during embryonic development in a tissue-specific manner, including the optic cup (retina) and the lens [2]. This article provides an analysis of Psf2 function during eye development in Xenopus laevis.
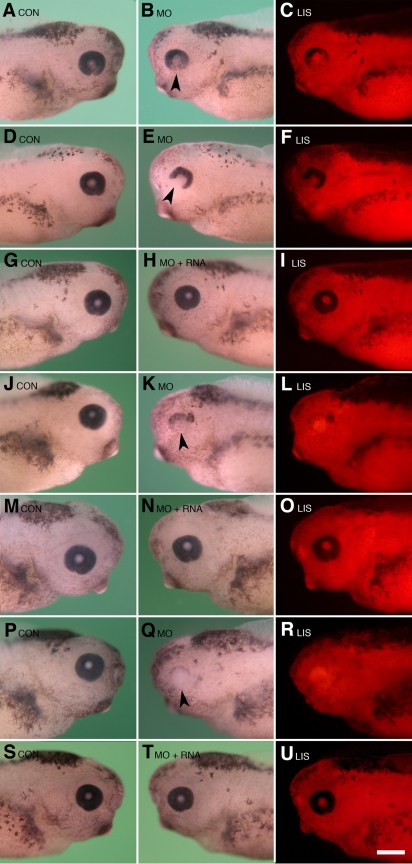
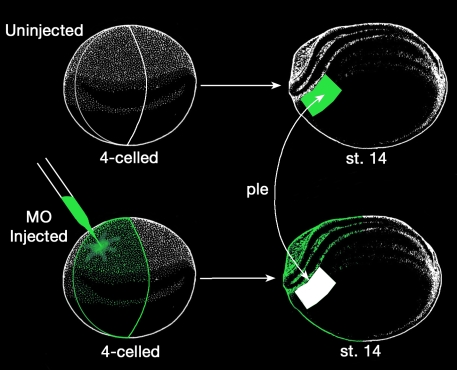
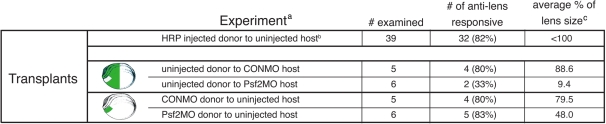
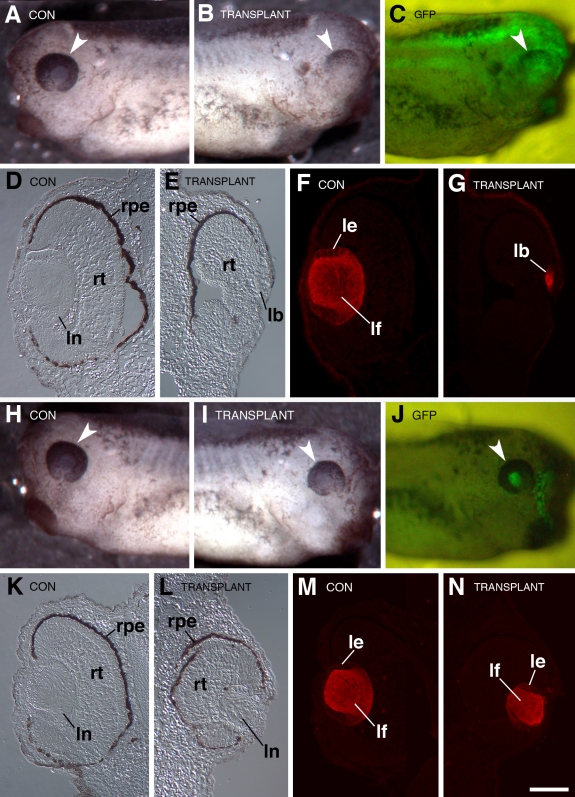
A morpholino targeted to Psf2 mRNA was designed to knockdown Psf2 translation and was injected into specific embryonic cells during early cleavage stages in the frog, Xenopus laevis. Injected embryos were assayed for specific defects in morphology, cell proliferation, and apoptosis. Synthetic Psf2 RNA was also co-injected with the morpholino to rescue morpholino-mediated developmental defects. It is well known that reciprocal inductive interactions control the development of the optic cup and lens. Therefore, control- and morpholino-injected embryos were used for reciprocal transplantation experiments to distinguish the intrinsic role of Psf2 in the development of the optic cup (retina) versus the lens.
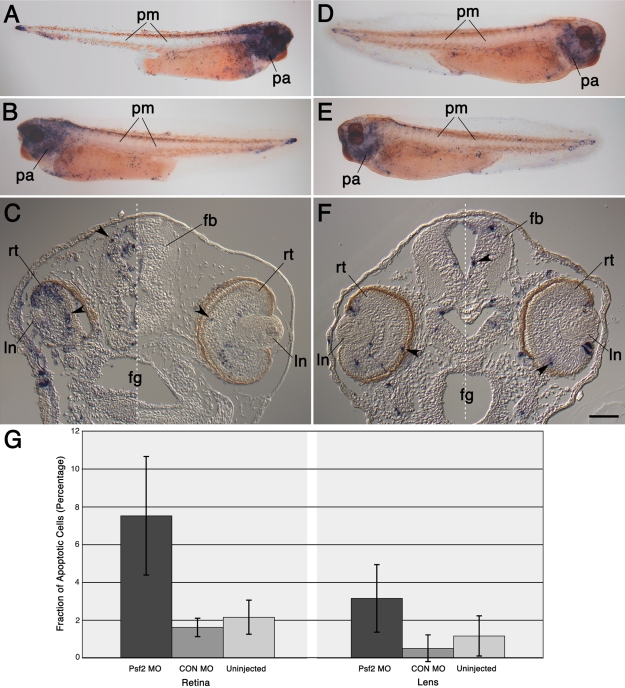
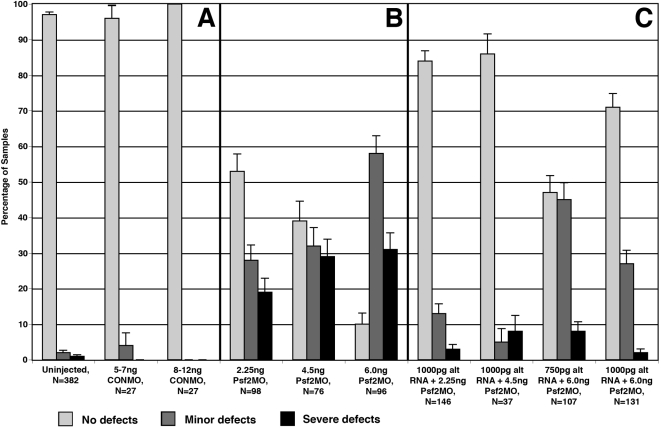
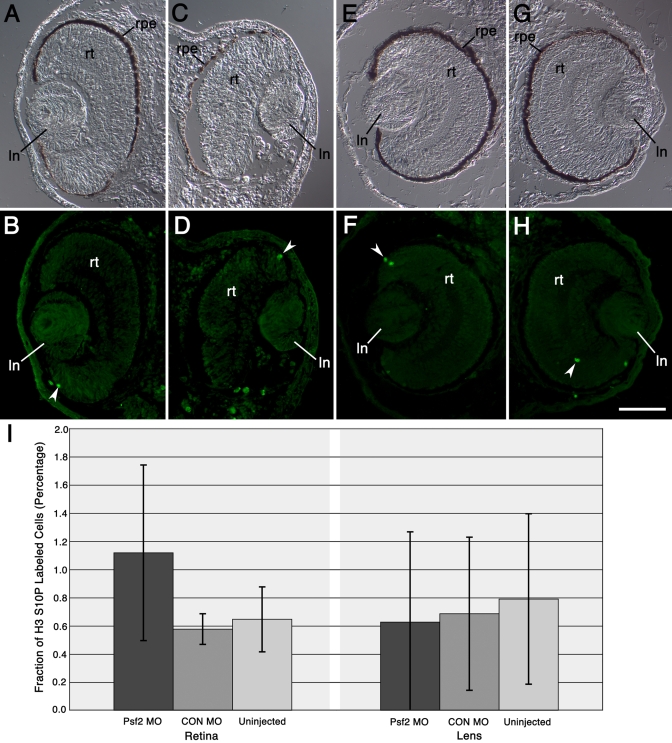
Morpholino-mediated knockdown of Psf2 expression resulted in dosage-dependent phenotypes, which included microphthalmia, incomplete closure of the ventral retinal fissure, and retinal and lens dysgenesis. Defects were also observed in other embryonic tissues that normally express Psf2 including the pharyngeal arches and the otic vesicle, although other tissues that express Psf2 were not found to be grossly defective. Eye defects could be rescued by co-injection of synthetic Psf2 RNA. Examination of cell proliferation via an antibody against phospho-histone H3 S10P revealed no significant differences in the retina and lens following Psf2 knockdown. However, there was a significant increase in the level of apoptosis in retinal as well as forebrain tissues, as revealed by TUNEL (terminal deoxynucleotide transferase dUTP nick end labeling) assay.
The results demonstrate intrinsic roles for Psf2 in both retinal and to a lesser extent, lens tissues. Observed lens defects can mainly be attributed to deficiencies in retinal development and consequently the late phase of lens induction, which involves instructive cues from the optic cup. Developmental defects were not observed in all tissues that express Psf2, which could be related to differences in the translation of Psf2 or redundant effects of related factors such as proliferating cell nuclear antigen (PCNA).
Psf2(Sld5 2的伙伴)是GINS(五、一、二、三)异源四聚体的成员之一[1],在DNA复制中作为“滑动夹”发挥作用。先前的原位杂交分析表明,Psf2在胚胎发育过程中以组织特异性方式表达,包括视杯(视网膜)和晶状体[2]。本文对非洲爪蟾眼睛发育过程中Psf2的功能进行了分析。
设计了一种靶向Psf2 mRNA的吗啉代寡核苷酸,以敲低Psf2的翻译,并在非洲爪蟾早期卵裂阶段注射到特定的胚胎细胞中。对注射后的胚胎进行形态、细胞增殖和凋亡方面的特定缺陷检测。还将合成的Psf2 RNA与吗啉代寡核苷酸共同注射,以挽救吗啉代寡核苷酸介导的发育缺陷。众所周知,相互诱导相互作用控制视杯和晶状体的发育。因此,将对照和注射了吗啉代寡核苷酸的胚胎用于相互移植实验,以区分Psf2在视杯(视网膜)与晶状体发育中的内在作用。
吗啉代寡核苷酸介导的Psf2表达敲低导致剂量依赖性表型,包括小眼症、视网膜腹侧裂未完全闭合以及视网膜和晶状体发育不全。在其他正常表达Psf2的胚胎组织中也观察到缺陷,包括咽弓和耳泡,尽管未发现其他表达Psf2的组织存在明显缺陷。通过共注射合成的Psf2 RNA可以挽救眼睛缺陷。通过抗磷酸化组蛋白H3 S10P抗体检测细胞增殖,发现敲低Psf2后视网膜和晶状体无显著差异。然而,TUNEL(末端脱氧核苷酸转移酶dUTP缺口末端标记)检测显示,视网膜和前脑组织中的凋亡水平显著增加。
结果表明Psf2在视网膜以及程度较轻的晶状体组织中具有内在作用。观察到的晶状体缺陷主要可归因于视网膜发育缺陷以及晶状体诱导后期的缺陷,这涉及视杯的诱导信号。并非在所有表达Psf2的组织中都观察到发育缺陷,这可能与Psf2的翻译差异或相关因子(如增殖细胞核抗原(PCNA))的冗余作用有关。