Nishikura K, Yoo C, Kim U, Murray J M, Estes P A, Cash F E, Liebhaber S A
Wistar Institute of Anatomy and Biology, Philadelphia, PA 19104.
EMBO J. 1991 Nov;10(11):3523-32. doi: 10.1002/j.1460-2075.1991.tb04916.x.
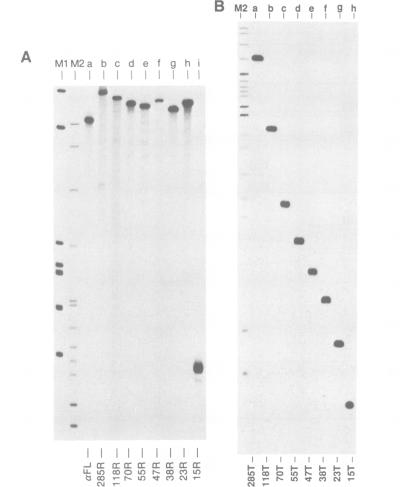
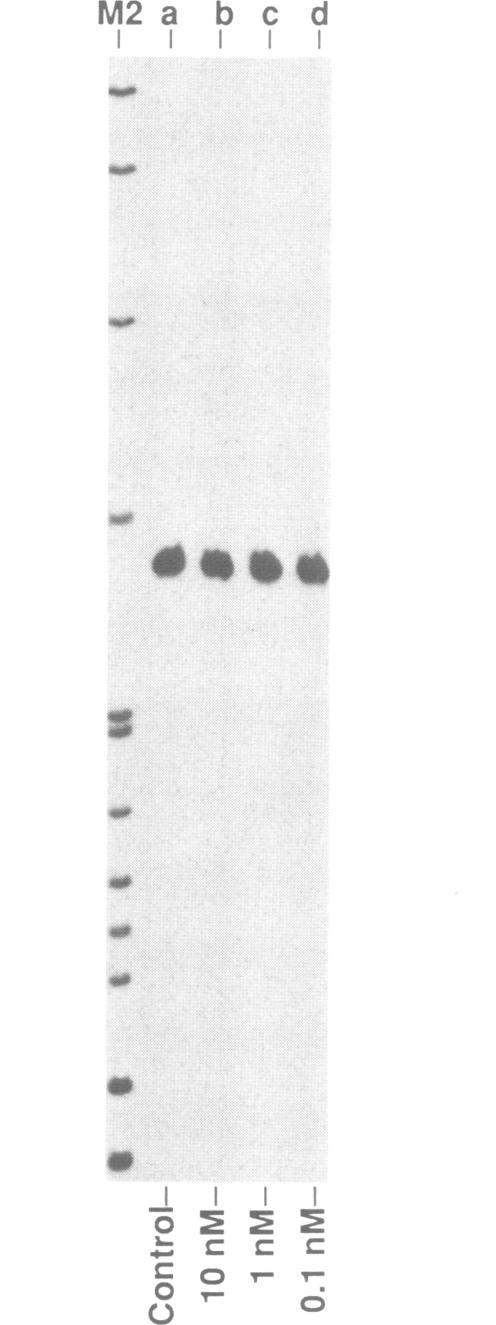
Double-stranded RNA (dsRNA) unwinding/modifying activity, which is present in a wide range of eukaryotic cells, has been previously shown to convert up to 50% of adenosine residues to inosines within intermolecular dsRNA. In the present study, we report that this activity also modifies, though slightly less efficiently, intramolecular double-stranded regions of synthetic RNAs. Our results widen the range of the possible biological substrates for the activity since many stem and loop type RNA secondary structures (intramolecular dsRNA), present in eukaryotic as well as viral transcripts, can potentially serve as substrates. In addition, we have found that the dsRNA unwinding/modifying activity requires a double-stranded region of at least 15-20 base pairs (bp) for substrate recognition. Furthermore, modification efficiency was found to be critically dependent on the length of the double-stranded region; as the size decreased below 100 bp, it dropped precipitously. Our results suggest that efficient modification may occur only with relatively long (greater than 100 bp) dsRNA, perhaps because multiple copies of the enzyme must be bound.
双链RNA(dsRNA)解旋/修饰活性存在于多种真核细胞中,此前研究表明,该活性可将分子间dsRNA中高达50%的腺苷残基转化为肌苷。在本研究中,我们报告称,这种活性也能修饰合成RNA的分子内双链区域,尽管效率略低。我们的研究结果拓宽了该活性可能作用的生物底物范围,因为真核生物以及病毒转录本中存在的许多茎环型RNA二级结构(分子内dsRNA)都有可能作为底物。此外,我们发现dsRNA解旋/修饰活性需要至少15 - 20个碱基对(bp)的双链区域来识别底物。此外,修饰效率被发现严重依赖于双链区域的长度;当大小降至100 bp以下时,修饰效率急剧下降。我们的研究结果表明,高效修饰可能仅发生在相对较长(大于100 bp)的dsRNA上,这可能是因为必须结合多个酶分子。