Department of Biology and University of Pennsylvania, Philadelphila, PA 19104-6018, USA.
Hum Mol Genet. 2009 Dec 15;18(24):4843-52. doi: 10.1093/hmg/ddp456. Epub 2009 Sep 25.
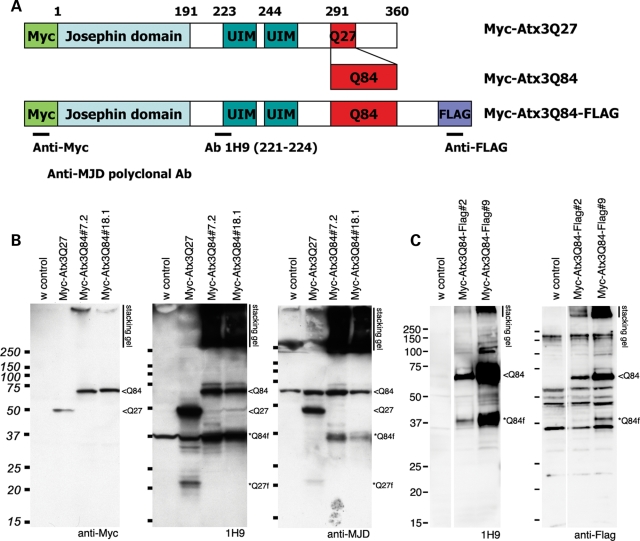
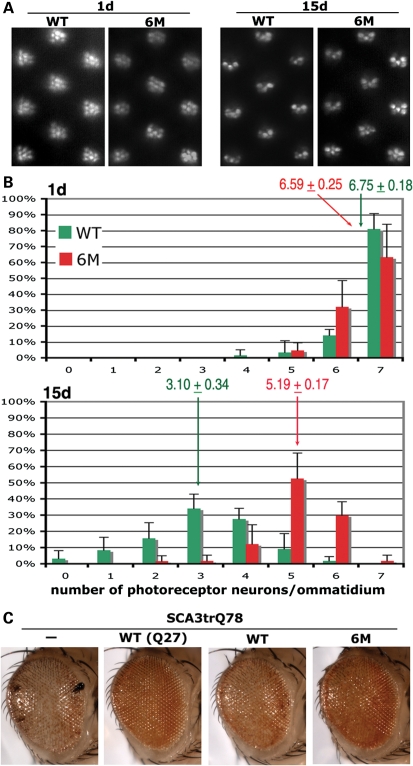
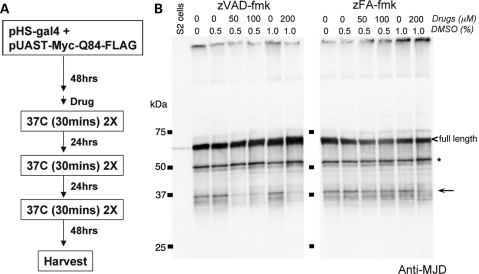
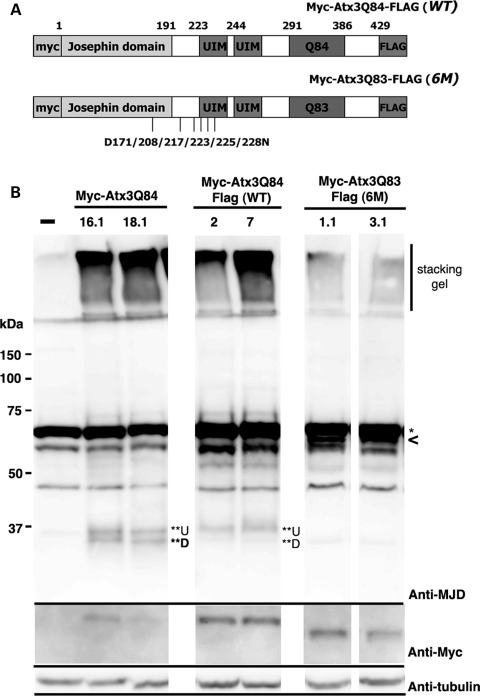
Protein cleavage is a common feature in human neurodegenerative disease. Ataxin-3 protein with an expanded polyglutamine (polyQ) repeat causes spinocerebellar ataxia type-3 (SCA3), also called Machado-Joseph disease, and is cleaved in mammalian cells, transgenic mice and SCA3 patient brain tissue. However, the pathological significance of Ataxin-3 cleavage has not been carefully examined. To gain insight into the significance of Ataxin-3 cleavage, we developed a Drosophila SL2 cell-based model as well as transgenic fly models. Our data indicate that Ataxin-3 protein cleavage is conserved in the fly and may be caspase-dependent as reported previously. Importantly, comparison of flies expressing either wild-type or caspase-site mutant proteins indicates that Ataxin-3 cleavage enhances neuronal loss in vivo. This genetic in vivo confirmation of the pathological role of Ataxin-3 cleavage indicates that therapies targeting Ataxin-3 cleavage might slow disease progression in SCA3 patients.
蛋白质裂解是人类神经退行性疾病的一个常见特征。具有扩增的多聚谷氨酰胺(polyQ)重复的 Ataxin-3 蛋白导致脊髓小脑共济失调 3 型(SCA3),也称为 Machado-Joseph 病,并在哺乳动物细胞、转基因小鼠和 SCA3 患者脑组织中被切割。然而,Ataxin-3 切割的病理意义尚未被仔细研究。为了深入了解 Ataxin-3 切割的意义,我们开发了一种基于果蝇 SL2 细胞的模型以及转基因果蝇模型。我们的数据表明,Ataxin-3 蛋白切割在果蝇中是保守的,并且可能像先前报道的那样依赖于半胱天冬酶。重要的是,比较表达野生型或半胱天冬酶切割位点突变蛋白的果蝇表明,Ataxin-3 切割增强了体内神经元的丢失。Ataxin-3 切割的这种遗传体内确认表明,针对 Ataxin-3 切割的治疗方法可能会减缓 SCA3 患者的疾病进展。