Department of Biochemistry and Molecular Biology and the Feist-Weiller Cancer Center, Louisiana State University Health Sciences Center, Shreveport, 71130, USA.
BMC Mol Biol. 2009 Dec 20;10:110. doi: 10.1186/1471-2199-10-110.
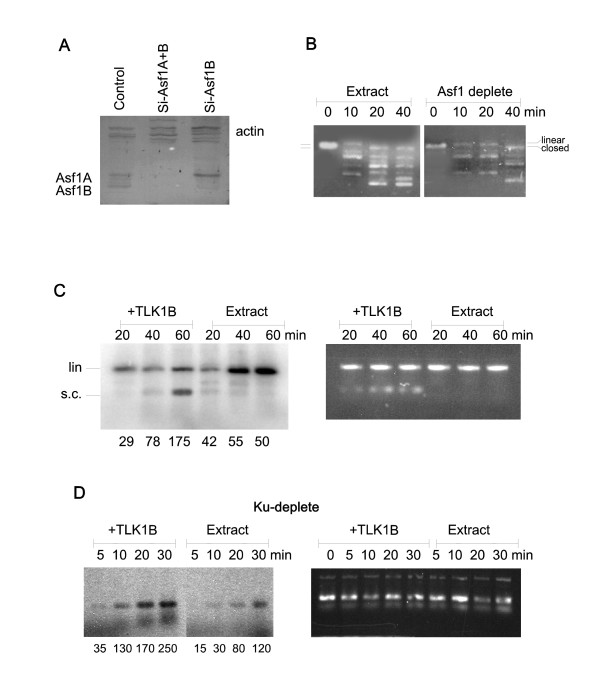
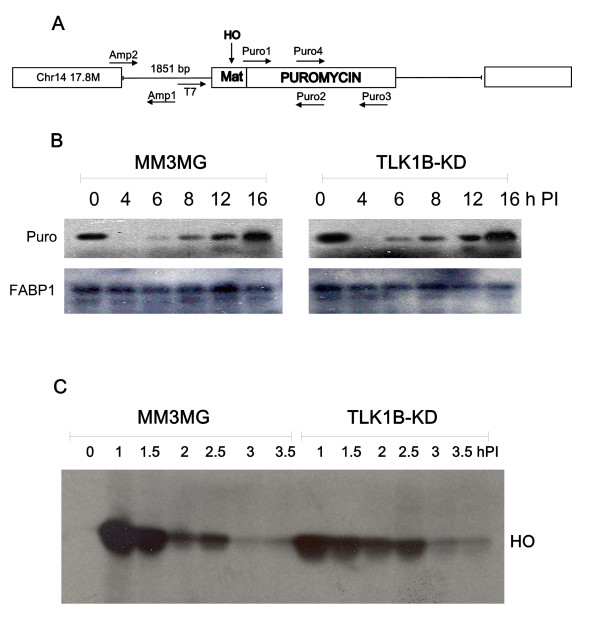
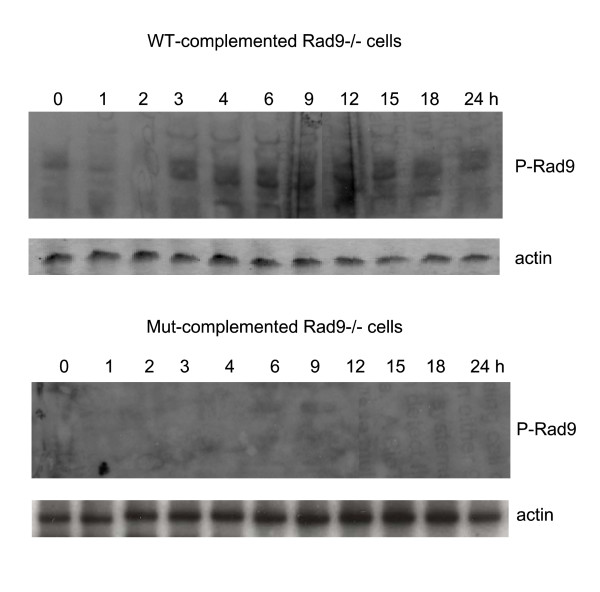
The Tousled-like kinases are involved in chromatin assembly, DNA repair, transcription, and chromosome segregation. Previous evidence indicated that TLK1B can promote repair of plasmids with cohesive ends in vitro, but it was inferred that the mechanism was indirect and via chromatin assembly, mediated by its interaction with the chromatin assembly factor Asf1. We recently identified Rad9 as a substrate of TLK1B, and we presented evidence that the TLK1B-Rad9 interaction plays some role in DSB repair. Hence the relative contribution of Asf1 and Rad9 to the protective effect of TLK1B in DSBs repair is not known. Using an adeno-HO-mediated cleavage system in MM3MG cells, we previously showed that overexpression of either TLK1B or a kinase-dead protein (KD) promoted repair and the assembly of Rad9 in proximity of the DSB at early time points post-infection. This established that it is a chaperone activity of TLK1B and not directly the kinase activity that promotes recruitment of 9-1-1 to the DSB. However, the phosphorylation of Rad9(S328) by TLK1B appeared important for mediating a cell cycle checkpoint, and thus, this phosphorylation of Rad9 may have other effects on 9-1-1 functionality.
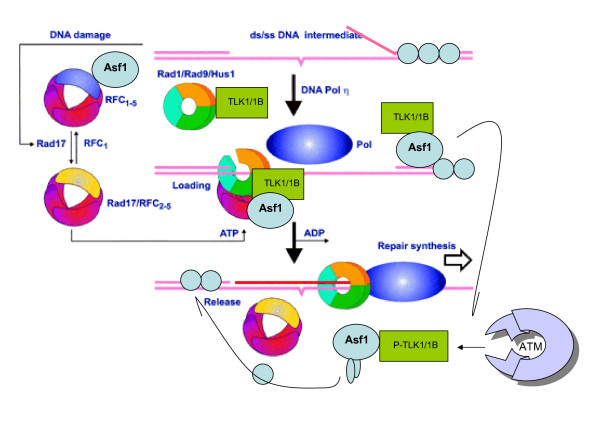
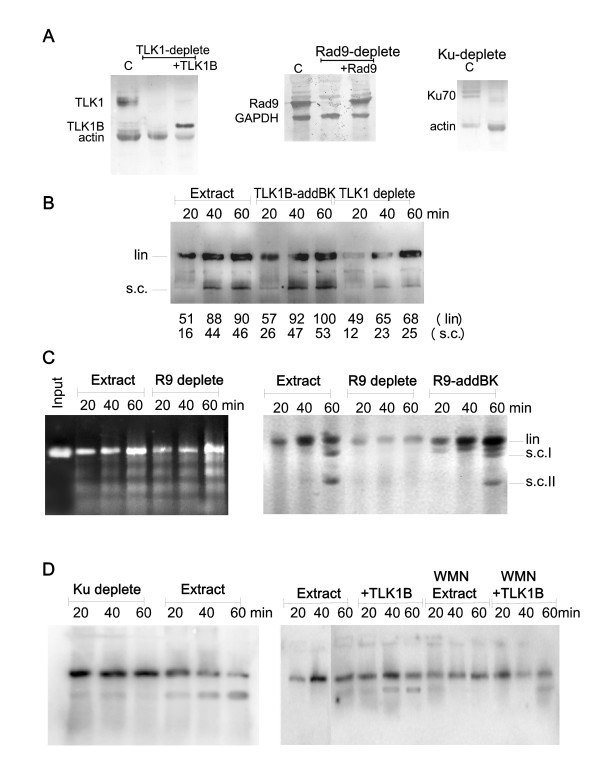
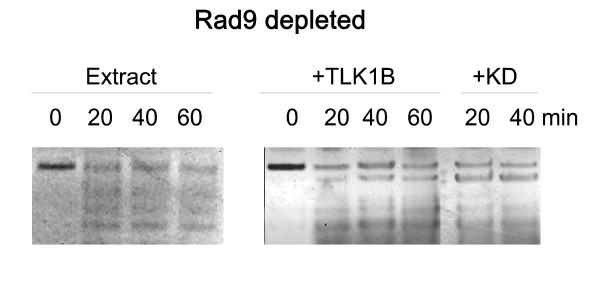
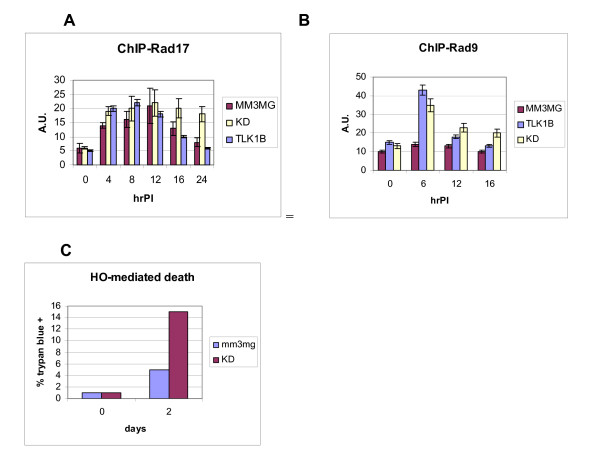
Here we present direct evidence that TLK1B can promote repair of linearized plasmids with incompatible ends that require processing prior to ligation. Immunodepletion of Rad9 indicated that Rad9 was important for processing the ends preceding ligation, suggesting that the interaction of TLK1B with Rad9 is a key mediator for this type of repair. Ligation of incompatible ends also required DNA-PK, as addition of wortmannin or immunodepletion of Ku70 abrogated ligation. Depletion of Ku70 prevented the ligation of the plasmid but did not affect stimulation of the fill-in of the ends by added TLK1B, which was attributed to Rad9. From experiments with the HO-cleavage system, we now show that Rad17, a subunit of the "clamp loader", associates normally with the DSB in KD-overexpressing cells. However, the subsequent release of Rad17 and Rad9 upon repair of the DSB was significantly slower in these cells compared to controls or cells expressing wt-TLK1B.
TLKs play important roles in DNA repair, not only by modulation of chromatin assembly via Asf1, but also by a more direct function in processing the ends of a DSB via interaction with Rad9. Inhibition of Rad9 phosphorylation in KD-overexpressing cells may have consequences in signaling completion of the repair and cell cycle re-entry, and could explain a loss of viability from DSBs in these cells.
Tousled 样激酶参与染色质组装、DNA 修复、转录和染色体分离。先前的证据表明,TLK1B 可以在体外促进带有粘性末端的质粒的修复,但据推断,这种机制是间接的,通过染色质组装,由其与染色质组装因子 Asf1 的相互作用介导。我们最近鉴定出 Rad9 是 TLK1B 的底物,我们提供的证据表明,TLK1B-Rad9 相互作用在 DSB 修复中发挥了一定的作用。因此,Asf1 和 Rad9 对 TLK1B 在 DSB 修复中的保护作用的相对贡献尚不清楚。使用 MM3MG 细胞中的腺病毒-HO 介导的切割系统,我们之前表明,TLK1B 或激酶失活蛋白(KD)的过表达均促进了感染后早期靠近 DSB 处的 Rad9 的修复和组装。这证明了它是 TLK1B 的伴侣活性,而不是直接的激酶活性,促进了 9-1-1 募集到 DSB。然而,TLK1B 对 Rad9(S328)的磷酸化似乎对介导细胞周期检查点很重要,因此,Rad9 的这种磷酸化可能对 9-1-1 的功能有其他影响。
这里我们提供了直接的证据,表明 TLK1B 可以促进具有不兼容末端的线性化质粒的修复,这些末端需要在连接前进行处理。Rad9 的免疫耗竭表明,Rad9 对于连接前的末端处理很重要,这表明 TLK1B 与 Rad9 的相互作用是这种类型修复的关键介质。不兼容末端的连接也需要 DNA-PK,因为添加wortmannin 或免疫耗竭 Ku70 会阻断连接。Ku70 的耗竭阻止了质粒的连接,但不影响添加的 TLK1B 对末端填充的刺激,这归因于 Rad9。通过 HO 切割系统的实验,我们现在表明,Rad17,“夹子加载器”的一个亚基,在 KD 过表达细胞中与 DSB 正常结合。然而,与对照细胞或表达 wt-TLK1B 的细胞相比,这些细胞中 DSB 修复后 Rad17 和 Rad9 的随后释放明显较慢。
TLKs 在 DNA 修复中发挥重要作用,不仅通过 Asf1 调节染色质组装,而且还通过与 Rad9 相互作用直接参与 DSB 末端的处理。在 KD 过表达细胞中抑制 Rad9 磷酸化可能会对修复完成和细胞周期重新进入的信号产生影响,并可以解释这些细胞中 DSB 导致的细胞活力丧失。