Department of Genetics and Development, Columbia University, New York, New York, USA.
PLoS Genet. 2010 Jan 15;6(1):e1000809. doi: 10.1371/journal.pgen.1000809.
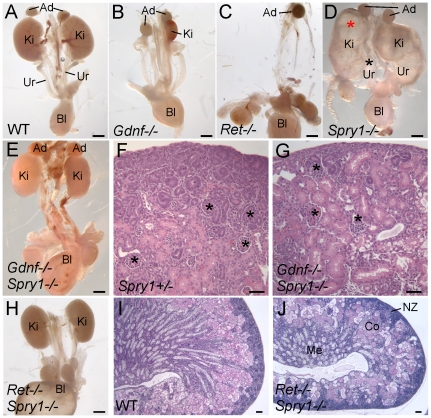
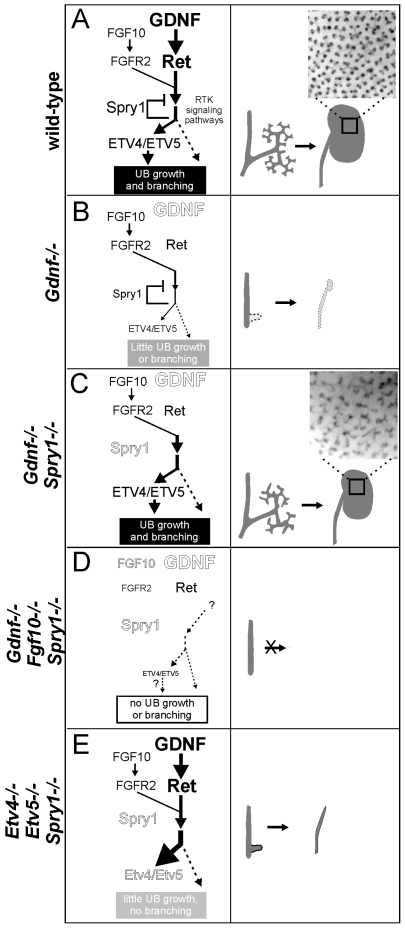
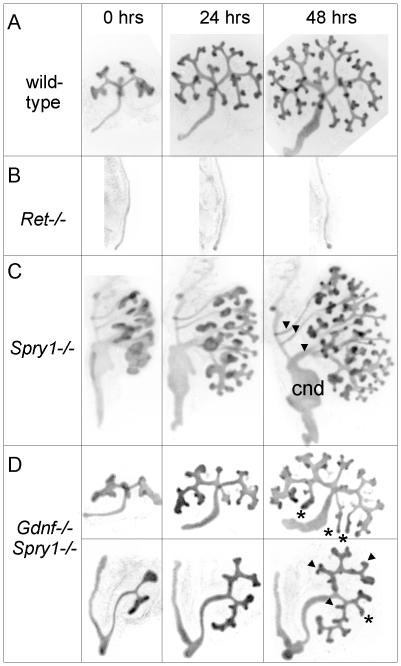
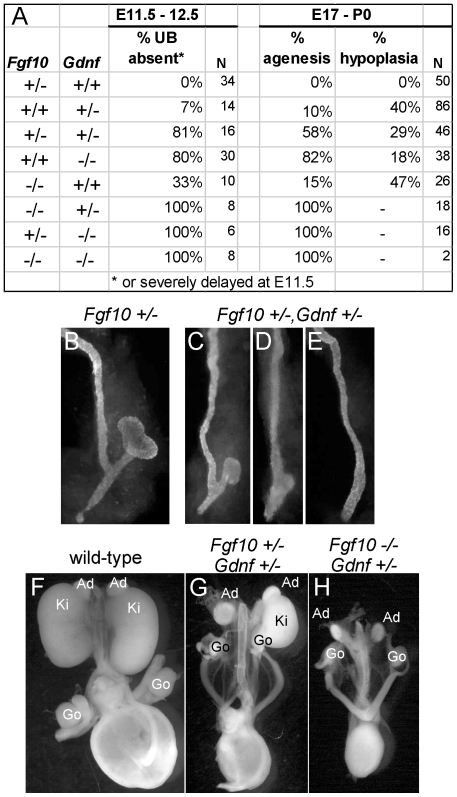
GDNF signaling through the Ret receptor tyrosine kinase (RTK) is required for ureteric bud (UB) branching morphogenesis during kidney development in mice and humans. Furthermore, many other mutant genes that cause renal agenesis exert their effects via the GDNF/RET pathway. Therefore, RET signaling is believed to play a central role in renal organogenesis. Here, we re-examine the extent to which the functions of Gdnf and Ret are unique, by seeking conditions in which a kidney can develop in their absence. We find that in the absence of the negative regulator Spry1, Gdnf, and Ret are no longer required for extensive kidney development. Gdnf-/-;Spry1-/- or Ret-/-;Spry1-/- double mutants develop large kidneys with normal ureters, highly branched collecting ducts, extensive nephrogenesis, and normal histoarchitecture. However, despite extensive branching, the UB displays alterations in branch spacing, angle, and frequency. UB branching in the absence of Gdnf and Spry1 requires Fgf10 (which normally plays a minor role), as removal of even one copy of Fgf10 in Gdnf-/-;Spry1-/- mutants causes a complete failure of ureter and kidney development. In contrast to Gdnf or Ret mutations, renal agenesis caused by concomitant lack of the transcription factors ETV4 and ETV5 is not rescued by removing Spry1, consistent with their role downstream of both RET and FGFRs. This shows that, for many aspects of renal development, the balance between positive signaling by RTKs and negative regulation of this signaling by SPRY1 is more critical than the specific role of GDNF. Other signals, including FGF10, can perform many of the functions of GDNF, when SPRY1 is absent. But GDNF/RET signaling has an apparently unique function in determining normal branching pattern. In contrast to GDNF or FGF10, Etv4 and Etv5 represent a critical node in the RTK signaling network that cannot by bypassed by reducing the negative regulation of upstream signals.
GDNF 信号通过 Ret 受体酪氨酸激酶(RTK)对于小鼠和人类肾脏发育过程中的输尿管芽(UB)分支形态发生是必需的。此外,许多其他导致肾发育不全的突变基因通过 GDNF/RET 途径发挥作用。因此,RET 信号被认为在肾脏发生中发挥核心作用。在这里,我们通过寻找在缺乏这些基因的情况下肾脏能够发育的条件,重新研究了 Gdnf 和 Ret 的功能的独特程度。我们发现,在缺乏负调控因子 Spry1 的情况下,Gdnf 和 Ret 不再是广泛肾脏发育所必需的。Gdnf-/-;Spry1-/-或 Ret-/-;Spry1-/-双突变体发育出具有正常输尿管的大型肾脏,具有高度分支的收集管,广泛的肾发生和正常的组织学结构。然而,尽管分支广泛,UB 显示出分支间距、角度和频率的改变。在没有 Gdnf 和 Spry1 的情况下,UB 的分支需要 Fgf10(其通常起次要作用),因为即使在 Gdnf-/-;Spry1-/-突变体中去除 Fgf10 的一个拷贝也会导致输尿管和肾脏发育完全失败。与 Gdnf 或 Ret 突变不同,同时缺乏转录因子 ETV4 和 ETV5 引起的肾发育不全不能通过去除 Spry1 来挽救,这与它们在 RET 和 FGFRs 下游的作用一致。这表明,对于肾脏发育的许多方面,RTK 的正信号和 SPRY1 对这种信号的负调节之间的平衡比 GDNF 的特定作用更关键。当 SPRY1 缺失时,其他信号,包括 FGF10,可以执行 GDNF 的许多功能。但是,GDNF/RET 信号在确定正常分支模式方面具有明显独特的功能。与 GDNF 或 FGF10 不同,Etv4 和 Etv5 代表 RTK 信号网络中的一个关键节点,不能通过减少上游信号的负调节来绕过。