Max F. Perutz Laboratories, Medical University of Vienna, Vienna Biocenter, Vienna, Austria.
PLoS Genet. 2010 Apr 29;6(4):e1000927. doi: 10.1371/journal.pgen.1000927.
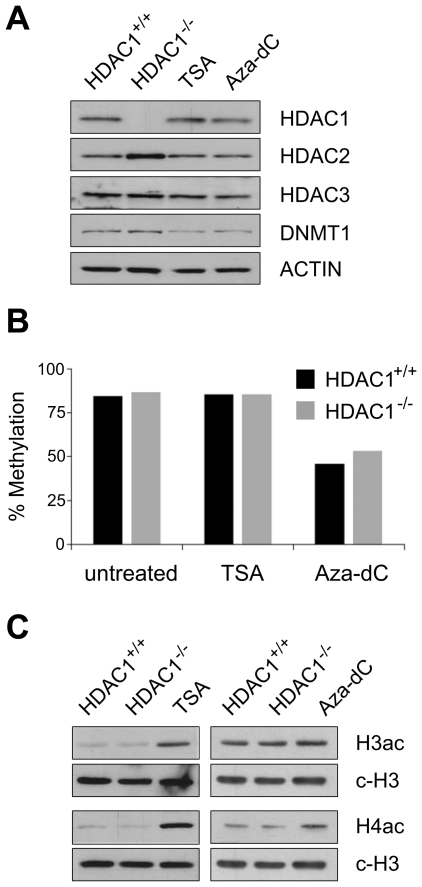
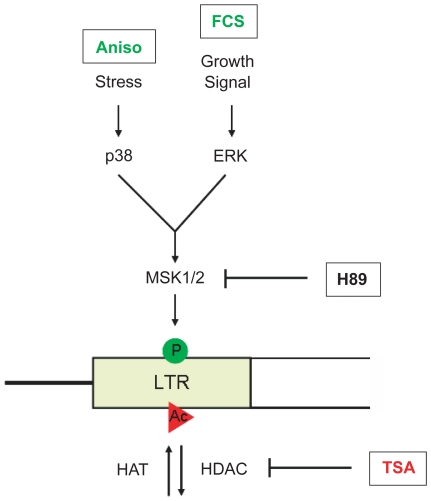
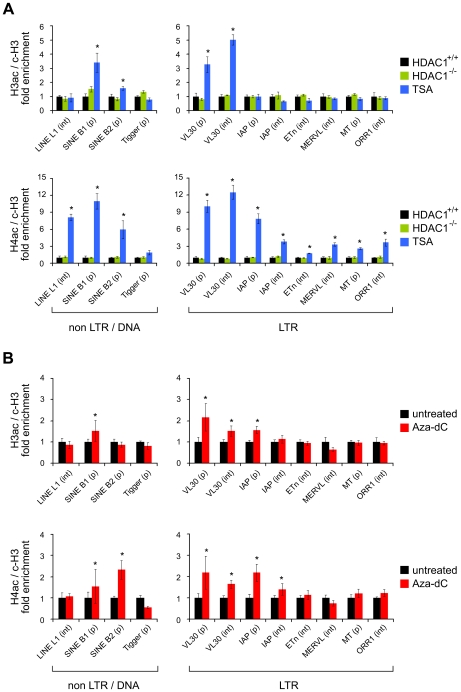
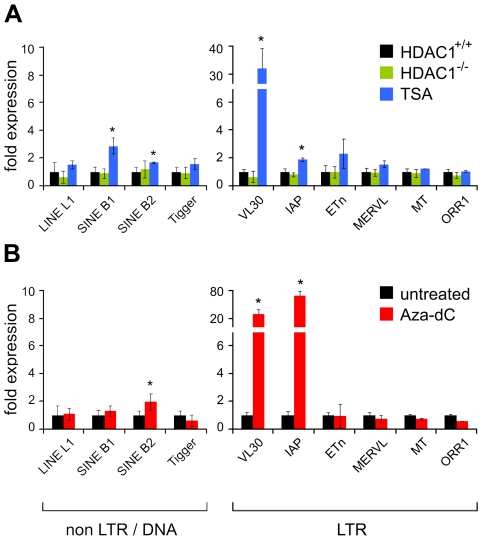
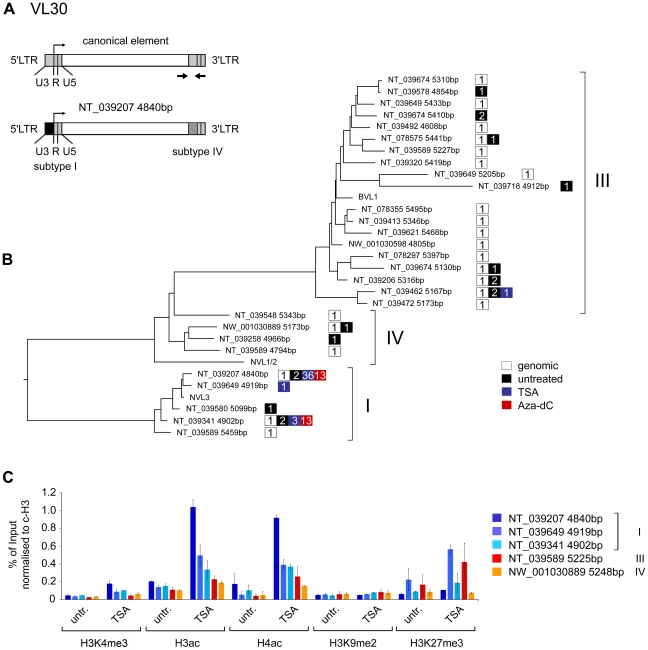
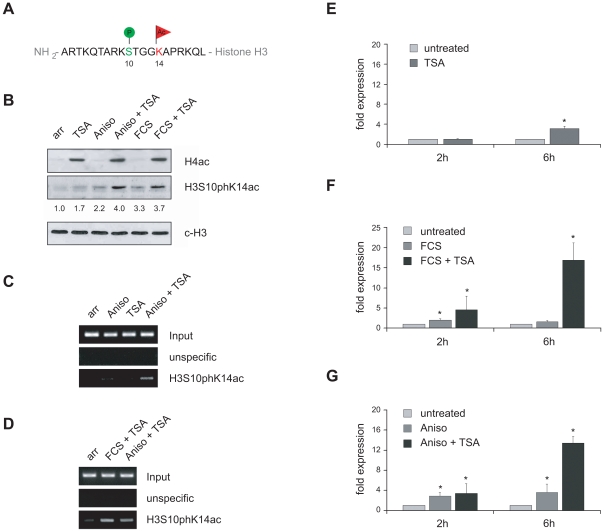
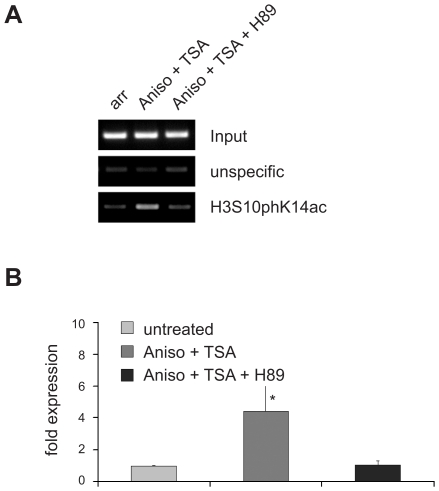
Large fractions of eukaryotic genomes contain repetitive sequences of which the vast majority is derived from transposable elements (TEs). In order to inactivate those potentially harmful elements, host organisms silence TEs via methylation of transposon DNA and packaging into chromatin associated with repressive histone marks. The contribution of individual histone modifications in this process is not completely resolved. Therefore, we aimed to define the role of reversible histone acetylation, a modification commonly associated with transcriptional activity, in transcriptional regulation of murine TEs. We surveyed histone acetylation patterns and expression levels of ten different murine TEs in mouse fibroblasts with altered histone acetylation levels, which was achieved via chemical HDAC inhibition with trichostatin A (TSA), or genetic inactivation of the major deacetylase HDAC1. We found that one LTR retrotransposon family encompassing virus-like 30S elements (VL30) showed significant histone H3 hyperacetylation and strong transcriptional activation in response to TSA treatment. Analysis of VL30 transcripts revealed that increased VL30 transcription is due to enhanced expression of a limited number of genomic elements, with one locus being particularly responsive to HDAC inhibition. Importantly, transcriptional induction of VL30 was entirely dependent on the activation of MAP kinase pathways, resulting in serine 10 phosphorylation at histone H3. Stimulation of MAP kinase cascades together with HDAC inhibition led to simultaneous phosphorylation and acetylation (phosphoacetylation) of histone H3 at the VL30 regulatory region. The presence of the phosphoacetylation mark at VL30 LTRs was linked with full transcriptional activation of the mobile element. Our data indicate that the activity of different TEs is controlled by distinct chromatin modifications. We show that activation of a specific mobile element is linked to a dual epigenetic mark and propose a model whereby phosphoacetylation of histone H3 is crucial for full transcriptional activation of VL30 elements.
真核生物基因组的大部分序列都含有重复序列,其中绝大多数来源于转座元件 (TEs)。为了使这些潜在有害的元件失活,宿主生物通过转座子 DNA 的甲基化和与抑制性组蛋白标记相关的染色质包装来沉默 TEs。在这个过程中,个别组蛋白修饰的贡献尚未完全确定。因此,我们旨在确定可逆组蛋白乙酰化在小鼠 TEs 的转录调控中的作用,这种修饰通常与转录活性相关。我们调查了在具有改变的组蛋白乙酰化水平的小鼠成纤维细胞中,十种不同的小鼠 TEs 的组蛋白乙酰化模式和表达水平,这些改变是通过用 Trichostatin A (TSA) 进行化学 HDAC 抑制或主要去乙酰化酶 HDAC1 的遗传失活来实现的。我们发现,一个包含病毒样 30S 元件 (VL30) 的 LTR 反转录转座子家族在 TSA 处理后表现出明显的组蛋白 H3 超乙酰化和强烈的转录激活。对 VL30 转录物的分析表明,VL30 转录的增加是由于少数基因组元件的表达增强,其中一个位点对 HDAC 抑制特别敏感。重要的是,VL30 的转录诱导完全依赖于 MAP 激酶途径的激活,导致组蛋白 H3 的丝氨酸 10 磷酸化。MAP 激酶级联的刺激加上 HDAC 抑制导致 VL30 调节区的组蛋白 H3 的同时磷酸化和乙酰化(磷酸乙酰化)。VL30 LTRs 上存在磷酸乙酰化标记与移动元件的完全转录激活有关。我们的数据表明,不同 TEs 的活性受不同的染色质修饰控制。我们表明,特定移动元件的激活与双重表观遗传标记有关,并提出了一个模型,即组蛋白 H3 的磷酸乙酰化对于 VL30 元件的完全转录激活至关重要。