Department of Biology and Rosenstiel Basic Medical Sciences Research Center, Brandeis University, Waltham, Massachusetts, United States of America.
PLoS Genet. 2010 May 27;6(5):e1000973. doi: 10.1371/journal.pgen.1000973.
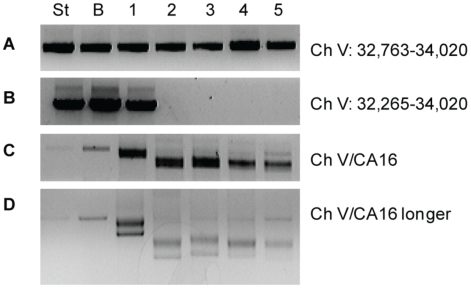
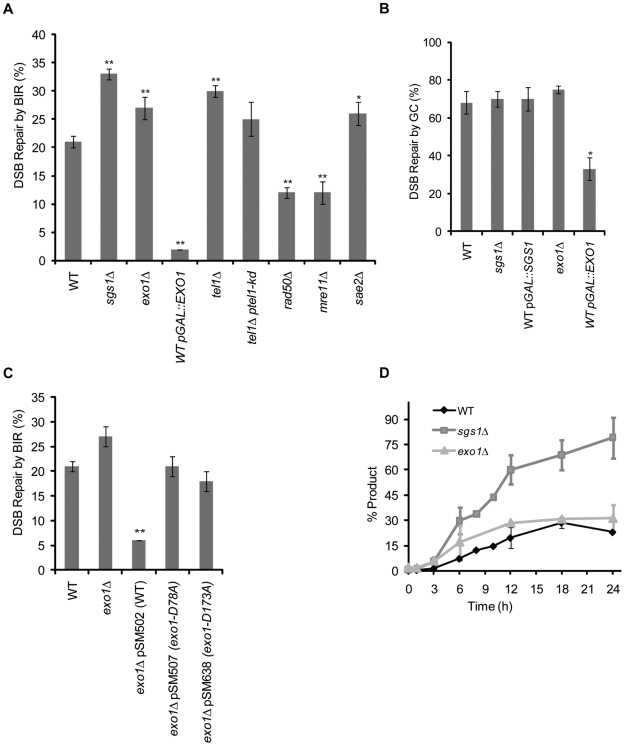
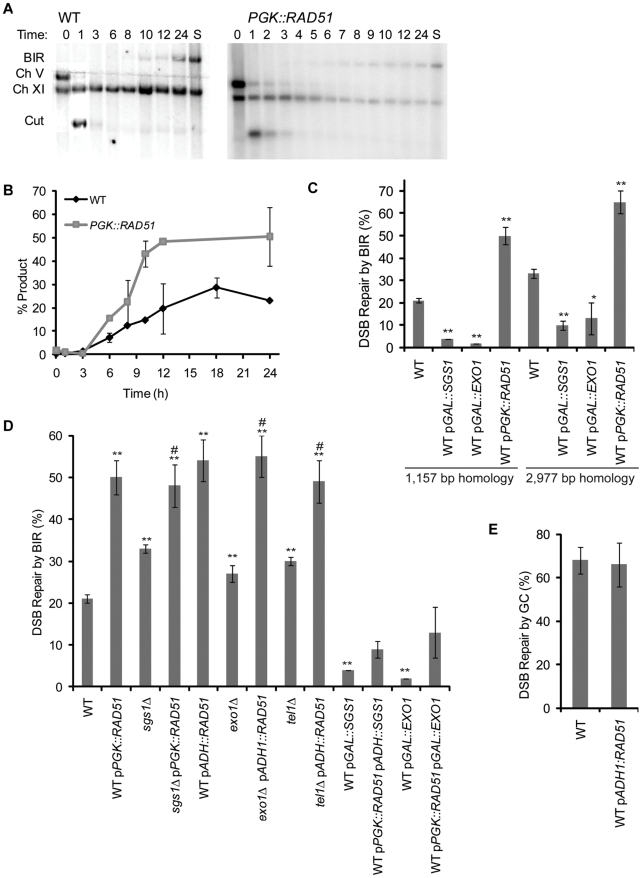
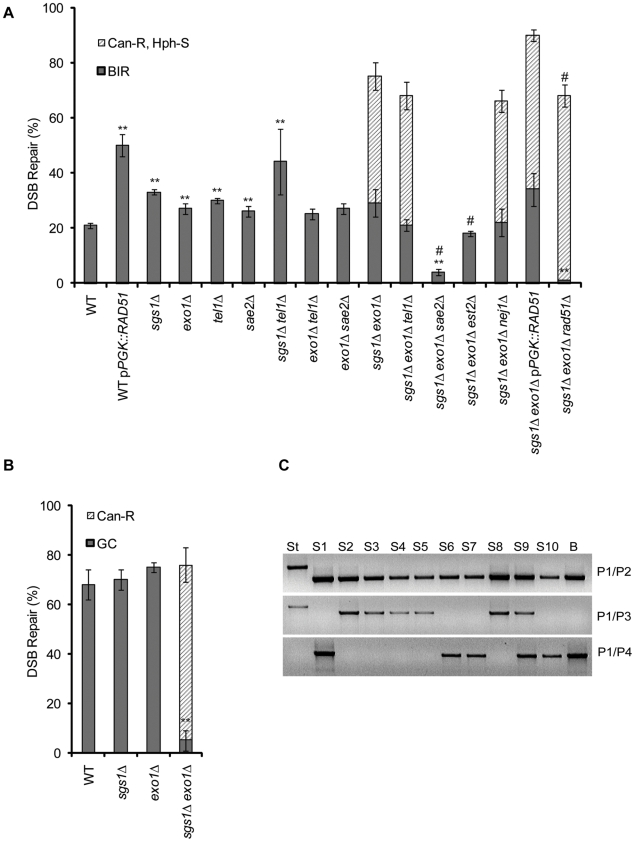
In budding yeast, an HO endonuclease-inducible double-strand break (DSB) is efficiently repaired by several homologous recombination (HR) pathways. In contrast to gene conversion (GC), where both ends of the DSB can recombine with the same template, break-induced replication (BIR) occurs when only the centromere-proximal end of the DSB can locate homologous sequences. Whereas GC results in a small patch of new DNA synthesis, BIR leads to a nonreciprocal translocation. The requirements for completing BIR are significantly different from those of GC, but both processes require 5' to 3' resection of DSB ends to create single-stranded DNA that leads to formation of a Rad51 filament required to initiate HR. Resection proceeds by two pathways dependent on Exo1 or the BLM homolog, Sgs1. We report that Exo1 and Sgs1 each inhibit BIR but have little effect on GC, while overexpression of either protein severely inhibits BIR. In contrast, overexpression of Rad51 markedly increases the efficiency of BIR, again with little effect on GC. In sgs1Delta exo1Delta strains, where there is little 5' to 3' resection, the level of BIR is not different from either single mutant; surprisingly, there is a two-fold increase in cell viability after HO induction whereby 40% of all cells survive by formation of a new telomere within a few kb of the site of DNA cleavage. De novo telomere addition is rare in wild-type, sgs1Delta, or exo1Delta cells. In sgs1Delta exo1Delta, repair by GC is severely inhibited, but cell viability remains high because of new telomere formation. These data suggest that the extensive 5' to 3' resection that occurs before the initiation of new DNA synthesis in BIR may prevent efficient maintenance of a Rad51 filament near the DSB end. The severe constraint on 5' to 3' resection, which also abrogates activation of the Mec1-dependent DNA damage checkpoint, permits an unprecedented level of new telomere addition.
在芽殖酵母中,HO 内切核酸酶诱导的双链断裂(DSB)可以通过几种同源重组(HR)途径有效地修复。与基因转换(GC)不同,GC 中 DSB 的两个末端都可以与同一个模板重组,而断裂诱导复制(BIR)仅发生在 DSB 的着丝粒近端末端能够定位同源序列时。GC 导致一小段新的 DNA 合成,而 BIR 导致非交互易位。完成 BIR 的要求与 GC 显著不同,但这两个过程都需要对 DSB 末端进行 5' 到 3' 切除,以产生单链 DNA,从而形成起始 HR 所需的 Rad51 丝。切除通过两条途径进行,依赖于 Exo1 或 BLM 同源物 Sgs1。我们报告说,Exo1 和 Sgs1 都抑制 BIR,但对 GC 影响很小,而两种蛋白质的过表达严重抑制 BIR。相反,Rad51 的过表达显著提高了 BIR 的效率,对 GC 的影响仍然很小。在 exo1Delta sgs1Delta 菌株中,由于几乎没有 5' 到 3' 切除,BIR 的水平与单个突变体没有区别;令人惊讶的是,在 HO 诱导后,细胞存活率提高了两倍,其中 40%的细胞通过在 DNA 切割位点附近几 kb 内形成新的端粒而存活。在野生型、sgs1Delta 或 exo1Delta 细胞中,新端粒的添加很少见。在 sgs1Delta exo1Delta 中,GC 的修复受到严重抑制,但由于新端粒的形成,细胞活力仍然很高。这些数据表明,在 BIR 中开始新的 DNA 合成之前发生的广泛的 5' 到 3' 切除可能会阻止 Rad51 丝在 DSB 末端附近的有效维持。对 5' 到 3' 切除的严格限制,也消除了 Mec1 依赖的 DNA 损伤检查点的激活,从而允许前所未有的新端粒添加水平。