CNRS UPR9073 (affiliated with University Paris 7-Denis Diderot), Institut de Biologie Physico-Chimique, 13 rue Pierre et Marie Curie, F-75005 Paris.
Nucleic Acids Res. 2011 Mar;39(5):1855-67. doi: 10.1093/nar/gkq1049. Epub 2010 Nov 8.
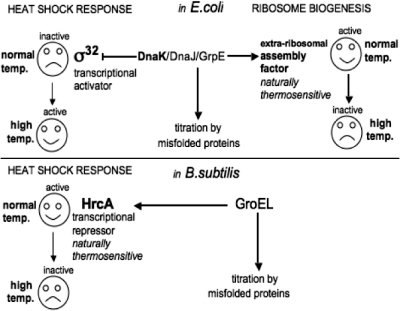
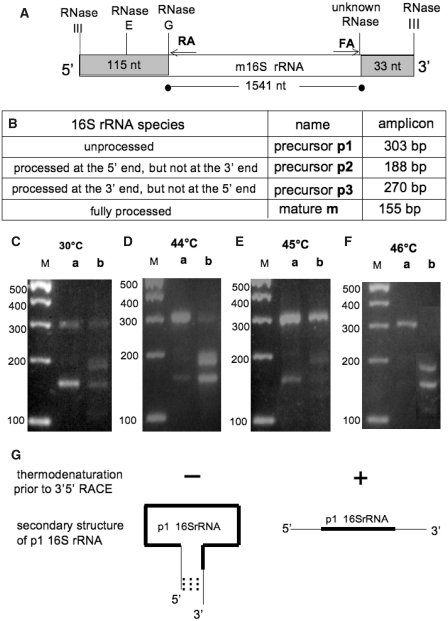
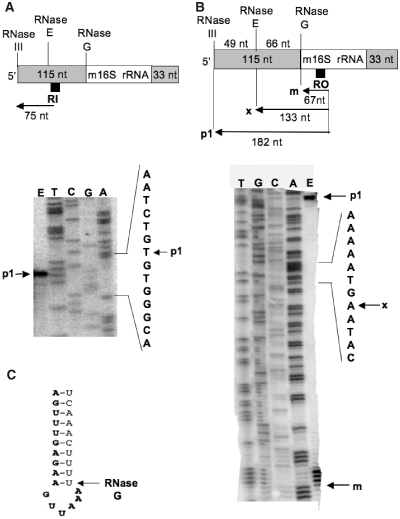
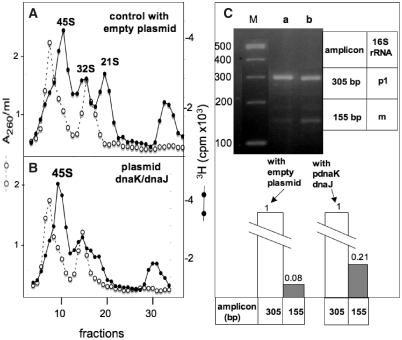
The late stages of 30S and 50S ribosomal subunits biogenesis have been studied in a wild-type (wt) strain of Escherichia coli (MC4100) subjected to a severe heat stress (45-46°C). The 32S and 45S ribosomal particles (precursors to 50S subunits) and 21S ribosomal particles (precursors to 30S subunits) accumulate under these conditions. They are authentic precursors, not degraded or dead-end particles. The 21S particles are shown, by way of a modified 3'5' RACE procedure, to contain 16S rRNA unprocessed, or processed at its 5' end, and not at the 3' end. This implies that maturation of 16S rRNA is ordered and starts at its 5'-terminus, and that the 3'-terminus is trimmed at a later step. This observation is not limited to heat stress conditions, but it also can be verified in bacteria growing at a normal temperature (30°C), supporting the idea that this is the general pathway. Assembly defects at very high temperature are partially compensated by plasmid-driven overexpression of the DnaK/DnaJ chaperones. The ribosome assembly pattern in wt bacteria under a severe heat stress is therefore reminiscent of that observed at lower temperatures in E. coli mutants lacking the chaperones DnaK or DnaJ.
在经历严重热应激(45-46°C)的大肠杆菌野生型(wt)菌株中,研究了 30S 和 50S 核糖体亚基生物发生的晚期阶段。在这些条件下,32S 和 45S 核糖体颗粒(50S 亚基的前体)和 21S 核糖体颗粒(30S 亚基的前体)积累。这些是真正的前体,不是降解或无出路的颗粒。通过改进的 3'5' RACE 程序,21S 颗粒显示出未加工的 16S rRNA 或在其 5' 端加工,而不是在 3' 端加工。这意味着 16S rRNA 的成熟是有序的,从其 5' 末端开始,而 3' 末端在稍后的步骤中被修剪。这一观察结果不仅限于热应激条件,而且在正常温度(30°C)下生长的细菌中也可以得到验证,支持了这是一般途径的观点。在非常高的温度下的组装缺陷部分通过质粒驱动的 DnaK/DnaJ 伴侣分子的过度表达得到补偿。因此,wt 细菌在严重热应激下的核糖体组装模式类似于在缺乏伴侣分子 DnaK 或 DnaJ 的大肠杆菌突变体中在较低温度下观察到的模式。