INSERM, UMR-S945, Université Pierre et Marie Curie (UPMC), Paris, France.
PLoS Pathog. 2011 May;7(5):e1002049. doi: 10.1371/journal.ppat.1002049. Epub 2011 May 12.
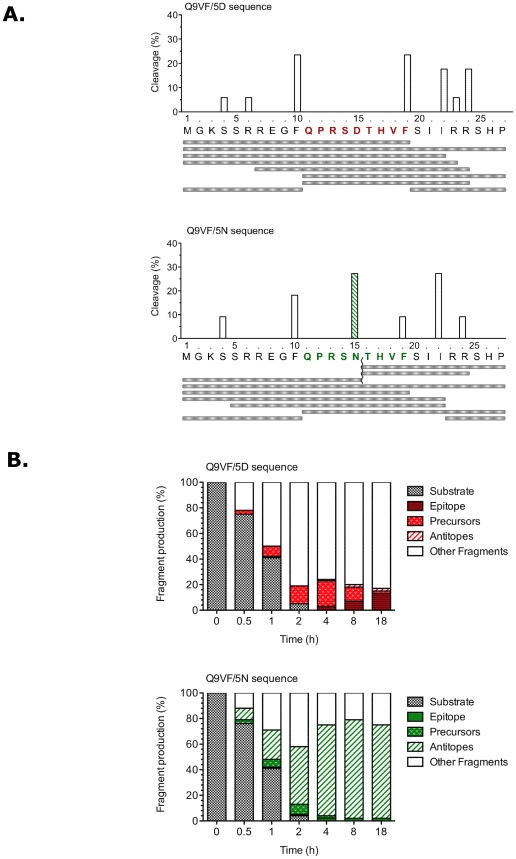
Cytotoxic CD8+ T cells (CTLs) play a critical role in controlling viral infections. HIV-infected individuals develop CTL responses against epitopes derived from viral proteins, but also against cryptic epitopes encoded by viral alternative reading frames (ARF). We studied here the mechanisms of HIV-1 escape from CTLs targeting one such cryptic epitope, Q9VF, encoded by an HIVgag ARF and presented by HLA-B07. Using PBMCs of HIV-infected patients, we first cloned and sequenced proviral DNA encoding for Q9VF. We identified several polymorphisms with a minority of proviruses encoding at position 5 an aspartic acid (Q9VF/5D) and a majority encoding an asparagine (Q9VF/5N). We compared the prevalence of each variant in PBMCs of HLA-B07+ and HLA-B07- patients. Proviruses encoding Q9VF/5D were significantly less represented in HLA-B07+ than in HLA-B07- patients, suggesting that Q9FV/5D encoding viruses might be under selective pressure in HLA-B07+ individuals. We thus analyzed ex vivo CTL responses directed against Q9VF/5D and Q9VF/5N. Around 16% of HLA-B*07+ patients exhibited CTL responses targeting Q9VF epitopes. The frequency and the magnitude of CTL responses induced with Q9VF/5D or Q9VF/5N peptides were almost equal indicating a possible cross-reactivity of the same CTLs on the two peptides. We then dissected the cellular mechanisms involved in the presentation of Q9VF variants. As expected, cells infected with HIV strains encoding for Q9VF/5D were recognized by Q9VF/5D-specific CTLs. In contrast, Q9VF/5N-encoding strains were neither recognized by Q9VF/5N- nor by Q9VF/5D-specific CTLs. Using in vitro proteasomal digestions and MS/MS analysis, we demonstrate that the 5N variation introduces a strong proteasomal cleavage site within the epitope, leading to a dramatic reduction of Q9VF epitope production. Our results strongly suggest that HIV-1 escapes CTL surveillance by introducing mutations leading to HIV ARF-epitope destruction by proteasomes.
细胞毒性 CD8+T 细胞(CTL)在控制病毒感染方面发挥着关键作用。HIV 感染者会针对源自病毒蛋白的表位产生 CTL 反应,但也会针对由病毒选择性阅读框(ARF)编码的隐匿表位产生反应。我们在这里研究了针对 HIV-1 逃避免疫监视的机制,该机制针对的是 HIVgag ARF 编码并由 HLA-B07 呈递的一个隐匿表位 Q9VF。我们使用 HIV 感染者的 PBMCs,首先克隆和测序了编码 Q9VF 的前病毒 DNA。我们发现了几个多态性,少数前病毒在第 5 位编码天冬氨酸(Q9VF/5D),而大多数前病毒编码天冬酰胺(Q9VF/5N)。我们比较了 HLA-B07+和 HLA-B07-患者 PBMC 中每种变体的流行率。编码 Q9VF/5D 的前病毒在 HLA-B07+患者中的代表性明显低于 HLA-B07-患者,表明 Q9FV/5D 编码的病毒可能在 HLA-B07+个体中受到选择压力。因此,我们分析了针对 Q9VF/5D 和 Q9VF/5N 的体外 CTL 反应。大约 16%的 HLA-B*07+患者表现出针对 Q9VF 表位的 CTL 反应。用 Q9VF/5D 或 Q9VF/5N 肽诱导的 CTL 反应的频率和幅度几乎相等,表明相同 CTL 对两种肽可能具有交叉反应性。然后,我们剖析了呈递 Q9VF 变体所涉及的细胞机制。正如预期的那样,感染编码 Q9VF/5D 的 HIV 株的细胞被 Q9VF/5D 特异性 CTL 识别。相反,Q9VF/5N 编码株既不能被 Q9VF/5N 特异性 CTL 识别,也不能被 Q9VF/5D 特异性 CTL 识别。通过体外蛋白酶体消化和 MS/MS 分析,我们证明 5N 变异在表位内引入了一个强的蛋白酶体切割位点,导致 Q9VF 表位产生急剧减少。我们的结果强烈表明,HIV-1 通过引入导致 ARF-表位被蛋白酶体破坏的突变来逃避 CTL 监视。