Centre for Integrative Physiology, University of Edinburgh, Edinburgh, United Kingdom.
PLoS One. 2011;6(6):e21056. doi: 10.1371/journal.pone.0021056. Epub 2011 Jun 9.
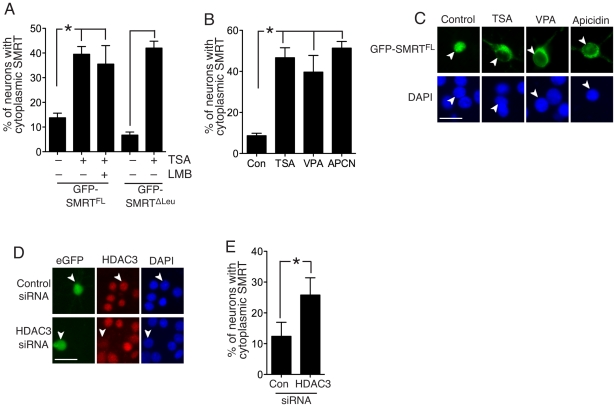
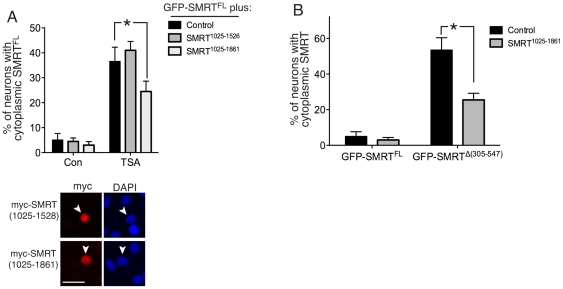
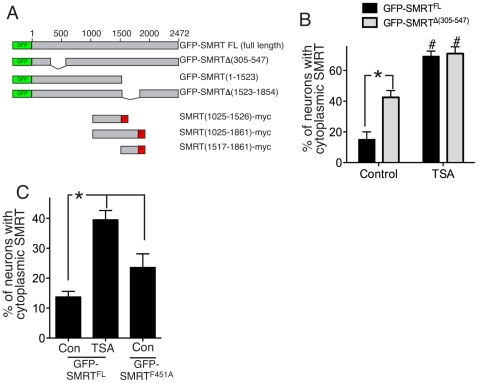
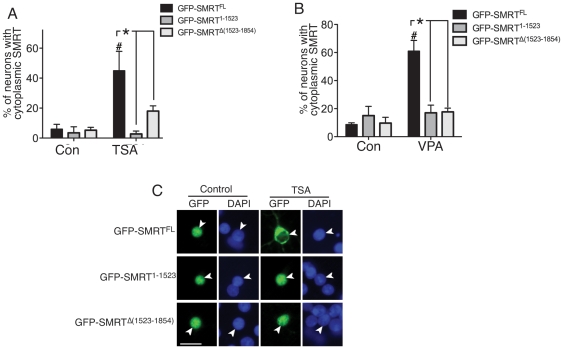
The transcriptional corepressor SMRT controls neuronal responsiveness of several transcription factors and can regulate neuroprotective and neurogenic pathways. SMRT is a multi-domain protein that complexes with HDAC3 as well as being capable of interactions with HDACs 1, 4, 5 and 7. We previously showed that in rat cortical neurons, nuclear localisation of SMRT requires histone deacetylase activity: Inhibition of class I/II HDACs by treatment with trichostatin A (TSA) causes redistribution of SMRT to the cytoplasm, and potentiates the activation of SMRT-repressed nuclear receptors. Here we have sought to identify the HDAC(s) and region(s) of SMRT responsible for anchoring it in the nucleus under normal circumstances and for mediating nuclear export following HDAC inhibition. We show that in rat cortical neurons SMRT export can be triggered by treatment with the class I-preferring HDAC inhibitor valproate and the HDAC2/3-selective inhibitor apicidin, and by HDAC3 knockdown, implicating HDAC3 activity as being required to maintain SMRT in the nucleus. HDAC3 interaction with SMRT's deacetylation activation domain (DAD) is known to be important for activation of HDAC3 deacetylase function. Consistent with a role for HDAC3 activity in promoting SMRT nuclear localization, we found that inactivation of SMRT's DAD by deletion or point mutation triggered partial redistribution of SMRT to the cytoplasm. We also investigated whether other regions of SMRT were involved in mediating nuclear export following HDAC inhibition. TSA- and valproate-induced SMRT export was strongly impaired by deletion of its repression domain-4 (RD4). Furthermore, over-expression of a region of SMRT containing the RD4 region suppressed TSA-induced export of full-length SMRT. Collectively these data support a model whereby SMRT's RD4 region can recruit factors capable of mediating nuclear export of SMRT, but whose function and/or recruitment is suppressed by HDAC3 activity. Furthermore, they underline the fact that HDAC inhibitors can cause reorganization and redistribution of corepressor complexes.
转录共抑制因子 SMRT 控制着几种转录因子的神经元反应性,并能调节神经保护和神经发生途径。SMRT 是一种多结构域蛋白,与 HDAC3 形成复合物,同时还能够与 HDAC1、4、5 和 7 相互作用。我们之前曾表明,在大鼠皮质神经元中,SMRT 的核定位需要组蛋白去乙酰化酶的活性:用曲古抑菌素 A(TSA)处理抑制 I/II 类 HDAC 会导致 SMRT 重新分布到细胞质中,并增强 SMRT 抑制的核受体的激活。在这里,我们试图确定在正常情况下将 SMRT 锚定在核内的 HDAC(s)和 SMRT 区域(s),以及在 HDAC 抑制后介导核输出的 HDAC(s)和 SMRT 区域(s)。我们发现在大鼠皮质神经元中,SMRT 出口可以通过用 I 类偏好的 HDAC 抑制剂丙戊酸和 HDAC2/3 选择性抑制剂 apicidin 处理以及通过 HDAC3 敲低来触发,这表明需要 HDAC3 活性来维持 SMRT 在核内。已知 HDAC3 与 SMRT 的去乙酰化激活结构域(DAD)的相互作用对于激活 HDAC3 去乙酰化酶功能很重要。与 HDAC3 活性在促进 SMRT 核定位中的作用一致,我们发现通过缺失或点突变使 SMRT 的 DAD 失活会触发 SMRT 部分重新分布到细胞质中。我们还研究了 SMRT 的其他区域是否参与介导 HDAC 抑制后的核输出。SMRT 的抑制结构域-4(RD4)缺失强烈削弱了 TSA 和丙戊酸诱导的 SMRT 出口。此外,含有 RD4 区域的 SMRT 区域的过表达抑制了 TSA 诱导的全长 SMRT 出口。这些数据共同支持一种模型,即 SMRT 的 RD4 区域可以募集能够介导 SMRT 核输出的因子,但 HDAC3 活性抑制了其功能和/或募集。此外,它们强调了这样一个事实,即 HDAC 抑制剂可以导致核心抑制复合物的重组和重分布。