HHMI and Kimmel Center for Biology and Medicine at Skirball Institute, Department of Cell Biology, New York University School of Medicine, New York, NY 10016, USA.
Development. 2011 Sep;138(18):4039-50. doi: 10.1242/dev.069187. Epub 2011 Aug 10.
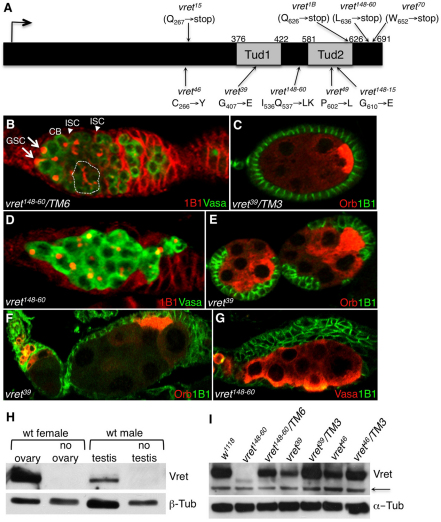
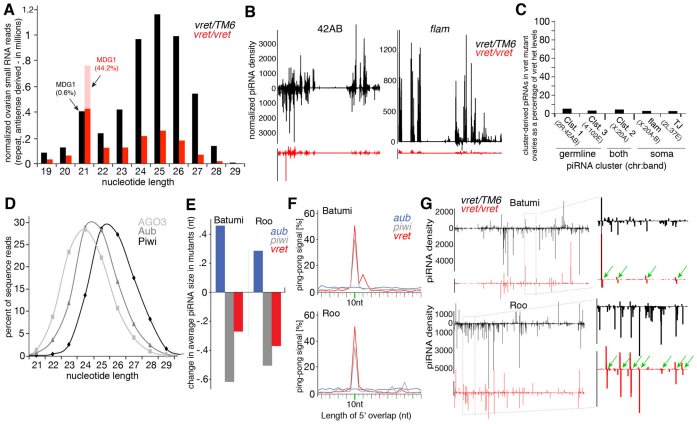
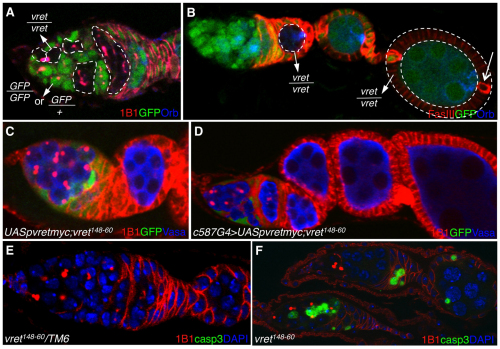
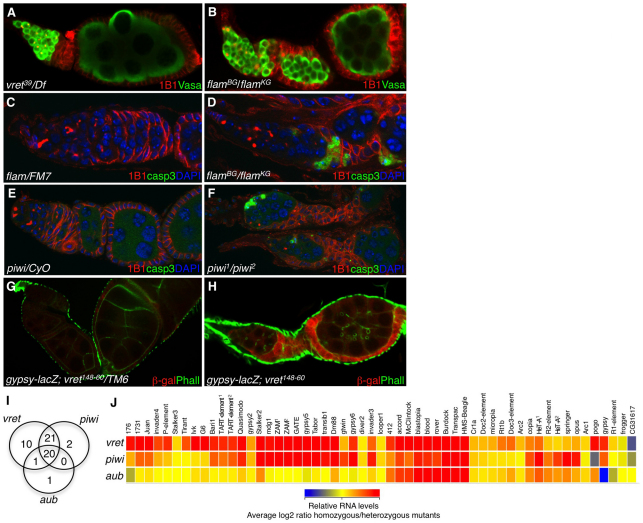
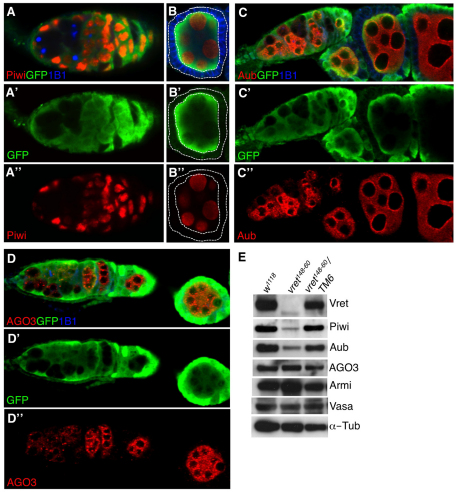
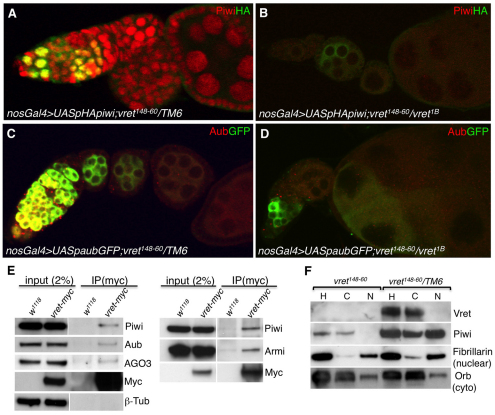
In Drosophila, Piwi proteins associate with Piwi-interacting RNAs (piRNAs) and protect the germline genome by silencing mobile genetic elements. This defense system acts in germline and gonadal somatic tissue to preserve germline development. Genetic control for these silencing pathways varies greatly between tissues of the gonad. Here, we identified Vreteno (Vret), a novel gonad-specific protein essential for germline development. Vret is required for piRNA-based transposon regulation in both germline and somatic gonadal tissues. We show that Vret, which contains Tudor domains, associates physically with Piwi and Aubergine (Aub), stabilizing these proteins via a gonad-specific mechanism that is absent in other fly tissues. In the absence of vret, Piwi-bound piRNAs are lost without changes in piRNA precursor transcript production, supporting a role for Vret in primary piRNA biogenesis. In the germline, piRNAs can engage in an Aub- and Argonaute 3 (AGO3)-dependent amplification in the absence of Vret, suggesting that Vret function can distinguish between primary piRNAs loaded into Piwi-Aub complexes and piRNAs engaged in the amplification cycle. We propose that Vret plays an essential role in transposon regulation at an early stage of primary piRNA processing.
在果蝇中,Piwi 蛋白与 Piwi 相互作用的 RNA(piRNAs)结合,通过沉默移动遗传元件来保护生殖系基因组。这种防御系统在生殖系和性腺体组织中发挥作用,以维持生殖系的发育。这些沉默途径的遗传控制在性腺的不同组织之间有很大的差异。在这里,我们鉴定了 Vreteno(Vret),一种新的性腺特异性蛋白,对生殖系发育是必需的。Vret 对于生殖系和体性腺组织中的基于 piRNA 的转座子调节是必需的。我们表明,Vret 含有 Tudor 结构域,与 Piwi 和 Aubergine(Aub)物理结合,通过一种在其他组织中不存在的性腺特异性机制稳定这些蛋白质。在 vret 缺失的情况下,Piwi 结合的 piRNAs 丢失,而 piRNA 前体转录物的产生没有变化,这支持了 Vret 在初级 piRNA 生物发生中的作用。在生殖系中,piRNAs 可以在没有 Vret 的情况下与 Aub 和 Argonaute 3(AGO3)依赖性扩增,这表明 Vret 功能可以区分加载到 Piwi-Aub 复合物中的初级 piRNAs 和参与扩增循环的 piRNAs。我们提出,Vret 在初级 piRNA 加工的早期阶段在转座子调节中发挥重要作用。