Department of Molecular Microbiology, John Innes Centre, Norwich Research Park, Norwich NR4 7UH, UK.
Nucleic Acids Res. 2011 Nov;39(21):9433-47. doi: 10.1093/nar/gkr640. Epub 2011 Aug 10.
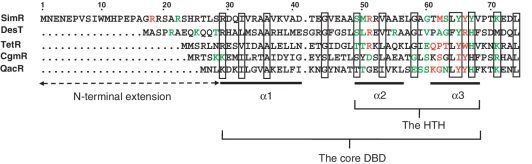
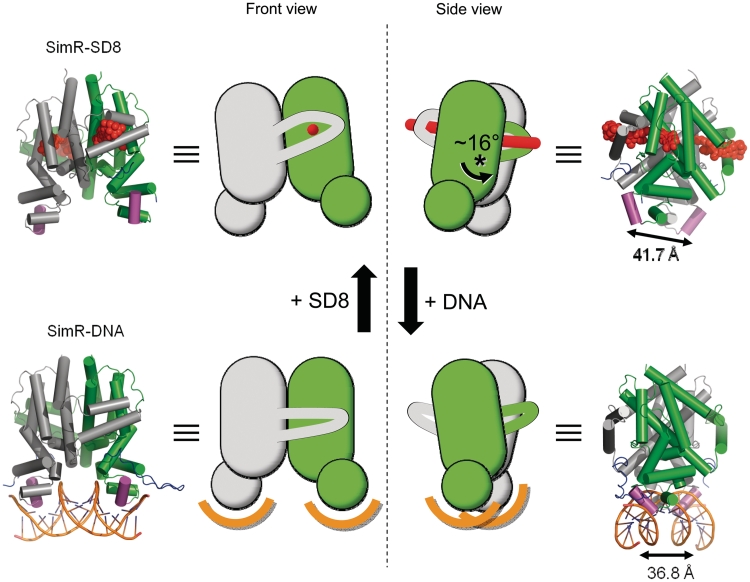
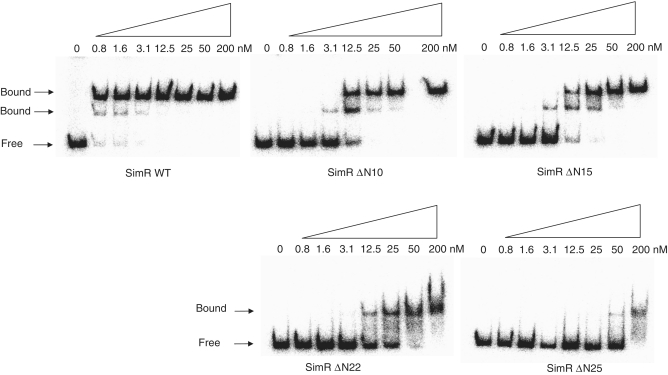
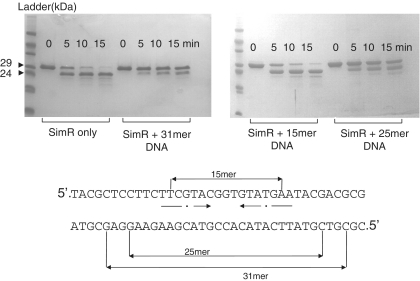
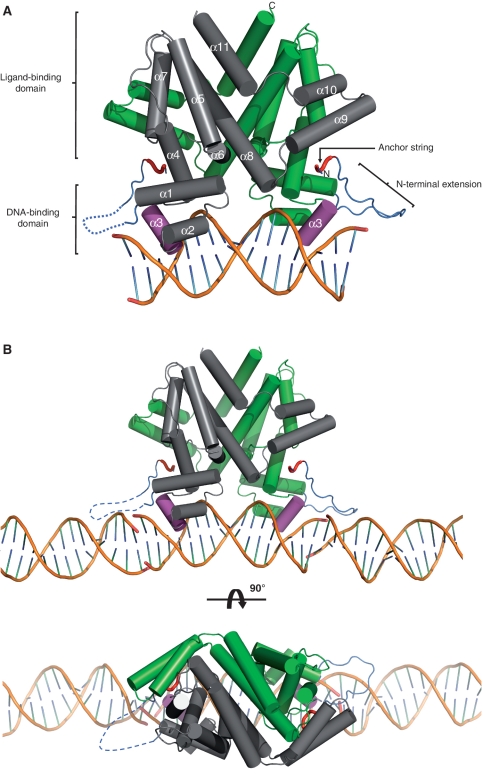
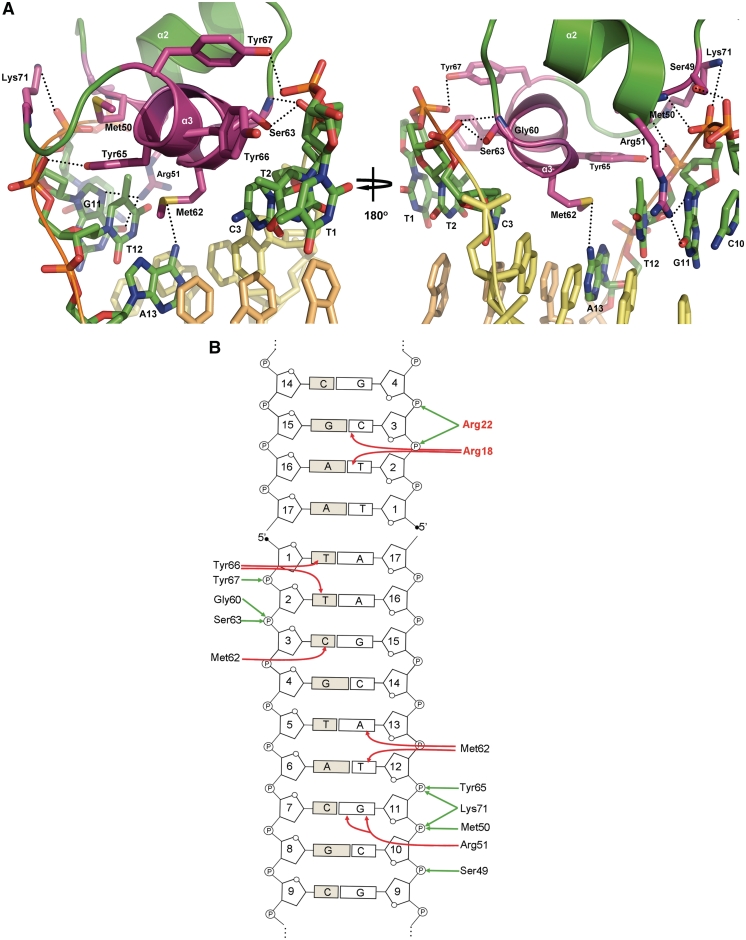
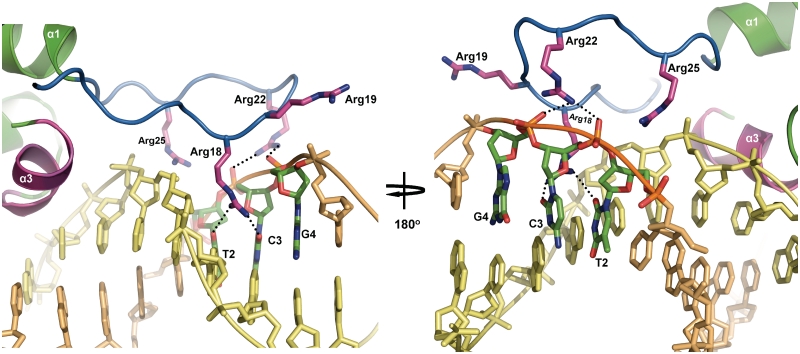
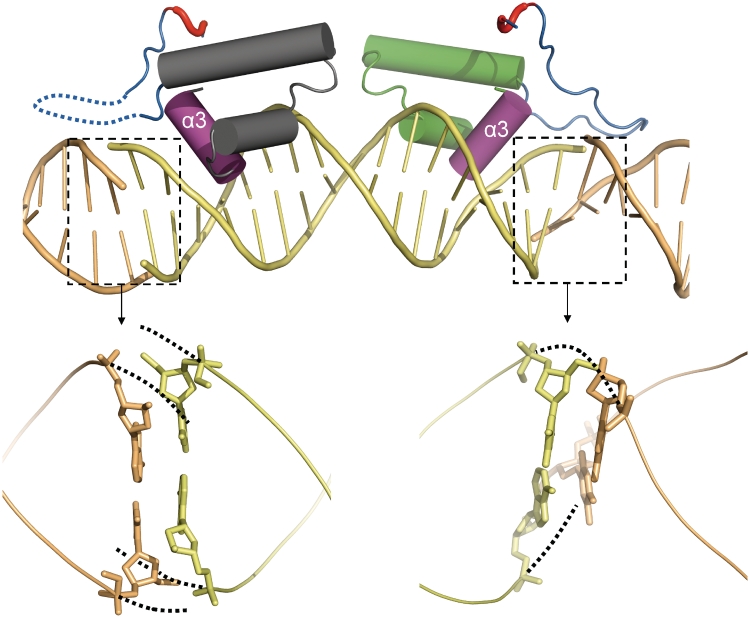
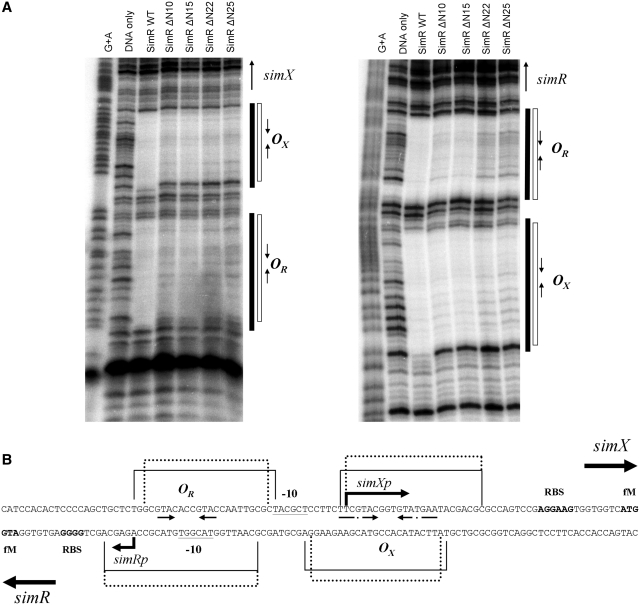
SimR, a TetR-family transcriptional regulator (TFR), controls the export of simocyclinone, a potent DNA gyrase inhibitor made by Streptomyces antibioticus. Simocyclinone is exported by a specific efflux pump, SimX and the transcription of simX is repressed by SimR, which binds to two operators in the simR-simX intergenic region. The DNA-binding domain of SimR has a classical helix-turn-helix motif, but it also carries an arginine-rich N-terminal extension. Previous structural studies showed that the N-terminal extension is disordered in the absence of DNA. Here, we show that the N-terminal extension is sensitive to protease cleavage, but becomes protease resistant upon binding DNA. We demonstrate by deletion analysis that the extension contributes to DNA binding, and describe the crystal structure of SimR bound to its operator sequence, revealing that the N-terminal extension binds in the minor groove. In addition, SimR makes a number of sequence-specific contacts to the major groove via its helix-turn-helix motif. Bioinformatic analysis shows that an N-terminal extension rich in positively charged residues is a feature of the majority of TFRs. Comparison of the SimR-DNA and SimR-simocyclinone complexes reveals that the conformational changes associated with ligand-mediated derepression result primarily from rigid-body rotation of the subunits about the dimer interface.
SimR 是 TetR 家族转录调节因子 (TFR),可控制来自链霉菌属放线菌的强力 DNA 拓扑异构酶抑制剂 simocyclinone 的外排。Simocyclinone 通过特定的外排泵 SimX 进行外排,而 SimX 的转录受到 SimR 的抑制,SimR 与 simR-simX 基因间区的两个操纵子结合。SimR 的 DNA 结合结构域具有经典的螺旋-转角-螺旋基序,但它还携带富含精氨酸的 N 端延伸。以前的结构研究表明,在没有 DNA 的情况下,N 端延伸是无序的。在这里,我们表明 N 端延伸对蛋白酶切割敏感,但与 DNA 结合后变得对蛋白酶具有抗性。我们通过缺失分析证明了延伸有助于 DNA 结合,并描述了与其操纵子序列结合的 SimR 的晶体结构,揭示了 N 端延伸在小沟中结合。此外,SimR 通过其螺旋-转角-螺旋基序与主沟进行了许多序列特异性接触。生物信息学分析表明,富含正电荷残基的 N 端延伸是大多数 TFR 的特征。SimR-DNA 和 SimR-simocyclinone 复合物的比较表明,与配体介导的去阻遏相关的构象变化主要来自于亚基围绕二聚体界面的刚体旋转。