Department of Medical Biochemistry and Biophysics, Umeå University, Umeå, Sweden.
Crit Rev Biochem Mol Biol. 2012 Jan-Feb;47(1):50-63. doi: 10.3109/10409238.2011.630372. Epub 2011 Nov 3.
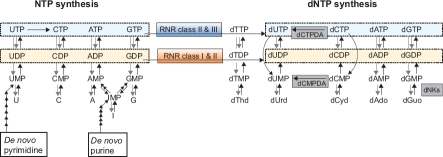
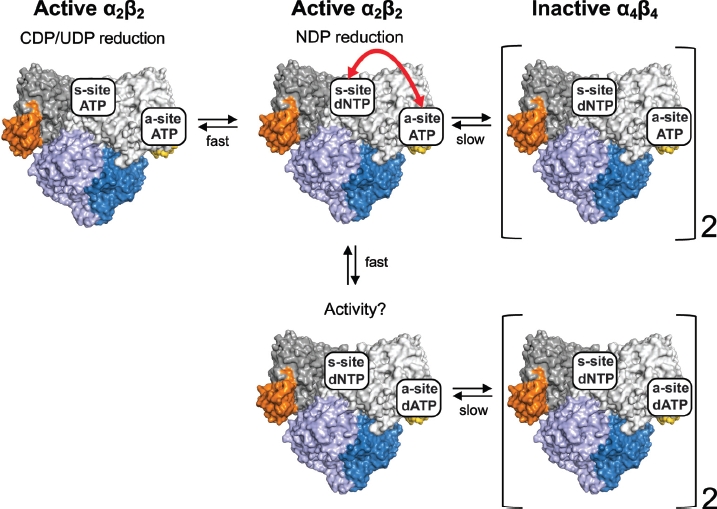
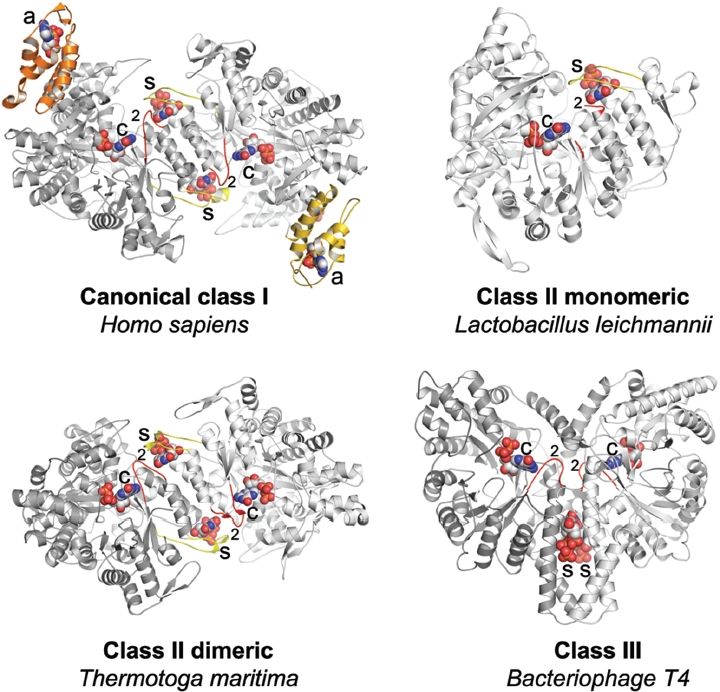
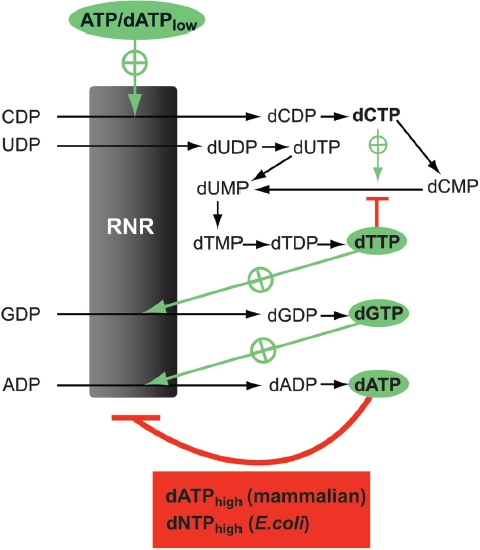
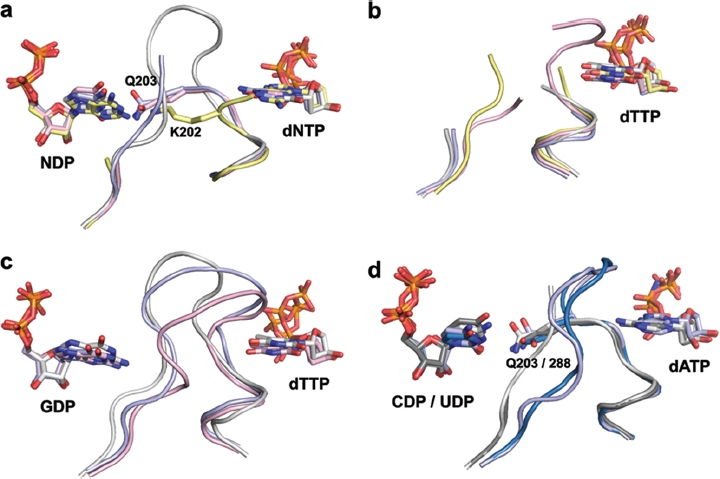
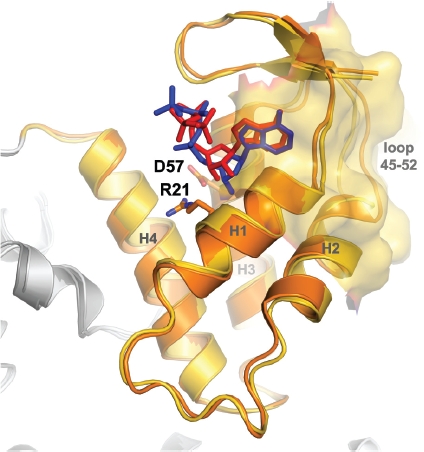
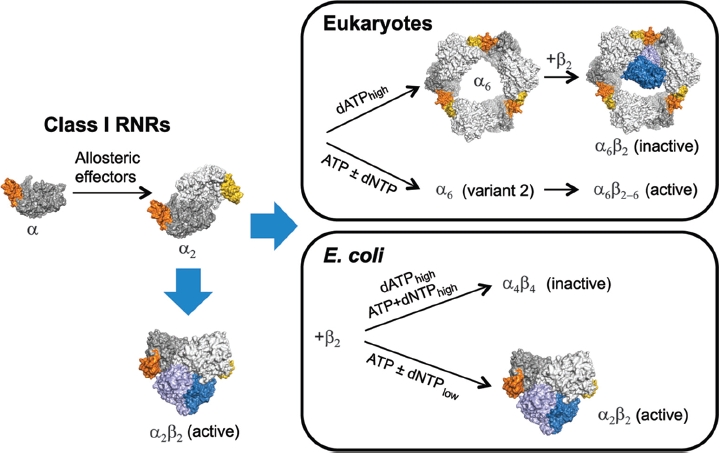
Ribonucleotide reductase (RNR) is the only source for de novo production of the four deoxyribonucleoside triphosphate (dNTP) building blocks needed for DNA synthesis and repair. It is crucial that these dNTP pools are carefully balanced, since mutation rates increase when dNTP levels are either unbalanced or elevated. RNR is the major player in this homeostasis, and with its four different substrates, four different allosteric effectors and two different effector binding sites, it has one of the most sophisticated allosteric regulations known today. In the past few years, the structures of RNRs from several bacteria, yeast and man have been determined in the presence of allosteric effectors and substrates, revealing new information about the mechanisms behind the allosteric regulation. A common theme for all studied RNRs is a flexible loop that mediates modulatory effects from the allosteric specificity site (s-site) to the catalytic site for discrimination between the four substrates. Much less is known about the allosteric activity site (a-site), which functions as an on-off switch for the enzyme's overall activity by binding ATP (activator) or dATP (inhibitor). The two nucleotides induce formation of different enzyme oligomers, and a recent structure of a dATP-inhibited α(6)β(2) complex from yeast suggested how its subunits interacted non-productively. Interestingly, the oligomers formed and the details of their allosteric regulation differ between eukaryotes and Escherichia coli. Nevertheless, these differences serve a common purpose in an essential enzyme whose allosteric regulation might date back to the era when the molecular mechanisms behind the central dogma evolved.
核糖核苷酸还原酶(RNR)是从头合成和修复 DNA 所需的四种脱氧核苷三磷酸(dNTP)构建块的唯一来源。这些 dNTP 池必须仔细平衡,因为当 dNTP 水平不平衡或升高时,突变率会增加。RNR 是这种动态平衡的主要参与者,它有四个不同的底物、四个不同的变构效应物和两个不同的效应物结合位点,因此具有当今已知的最复杂的变构调节之一。在过去的几年中,已经确定了几种细菌、酵母和人类的 RNR 结构,这些结构存在变构效应物和底物,揭示了变构调节背后机制的新信息。所有研究过的 RNR 都有一个共同的主题,即一个柔性环,它介导变构特异性位点(s-位点)到催化位点的调节效应,用于区分四个底物。关于变构活性位点(a-位点)的了解要少得多,该位点通过结合 ATP(激活剂)或 dATP(抑制剂)作为酶整体活性的开/关开关。这两个核苷酸诱导不同酶寡聚体的形成,最近来自酵母的 dATP 抑制的α(6)β(2)复合物的结构表明了其亚基如何非生产性地相互作用。有趣的是,形成的寡聚体及其变构调节的细节在真核生物和大肠杆菌之间存在差异。然而,这些差异在一种基本酶的变构调节中服务于共同的目的,这种酶的变构调节可能可以追溯到中央法则背后的分子机制演变的时代。