Laboratory of Cold Microbiology - Signals and Micro-Environment, UPRES EA 4312, University of Rouen, 55 rue Saint Germain, 27000 Evreux, France.
Sensors (Basel). 2009;9(9):6967-90. doi: 10.3390/s90906967. Epub 2009 Sep 2.
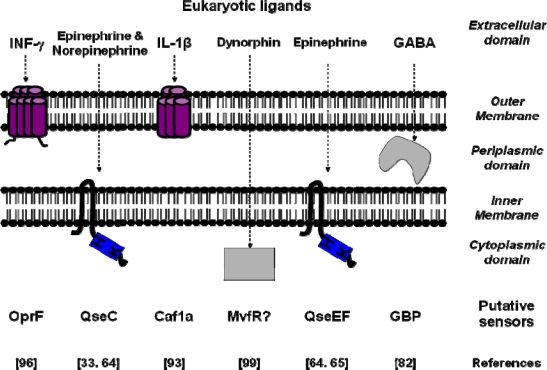
Ample evidence exists showing that eukaryotic signal molecules synthesized and released by the host can activate the virulence of opportunistic pathogens. The sensitivity of prokaryotes to host signal molecules requires the presence of bacterial sensors. These prokaryotic sensors, or receptors, have a double function: stereospecific recognition in a complex environment and transduction of the message in order to initiate bacterial physiological modifications. As messengers are generally unable to freely cross the bacterial membrane, they require either the presence of sensors anchored in the membrane or transporters allowing direct recognition inside the bacterial cytoplasm. Since the discovery of quorum sensing, it was established that the production of virulence factors by bacteria is tightly growth-phase regulated. It is now obvious that expression of bacterial virulence is also controlled by detection of the eukaryotic messengers released in the micro-environment as endocrine or neuro-endocrine modulators. In the presence of host physiological stress many eukaryotic factors are released and detected by Gram-negative bacteria which in return rapidly adapt their physiology. For instance, Pseudomonas aeruginosa can bind elements of the host immune system such as interferon-γ and dynorphin and then through quorum sensing circuitry enhance its virulence. Escherichia coli sensitivity to the neurohormones of the catecholamines family appears relayed by a recently identified bacterial adrenergic receptor. In the present review, we will describe the mechanisms by which various eukaryotic signal molecules produced by host may activate Gram-negative bacteria virulence. Particular attention will be paid to Pseudomonas, a genus whose representative species, P. aeruginosa, is a common opportunistic pathogen. The discussion will be particularly focused on the pivotal role played by these new types of pathogen sensors from the sensing to the transduction mechanism involved in virulence factors regulation. Finally, we will discuss the consequence of the impact of host signal molecules on commensally or opportunistic pathogens associated with different human tissue.
有充分的证据表明,宿主合成并释放的真核信号分子可以激活机会性病原体的毒力。原核生物对宿主信号分子的敏感性需要细菌传感器的存在。这些原核传感器或受体具有双重功能:在复杂环境中进行立体特异性识别,并传递信息以启动细菌生理修饰。由于信使通常无法自由穿过细菌膜,它们需要要么存在锚定在膜中的传感器,要么存在允许在细菌细胞质内直接识别的转运体。自从发现群体感应以来,人们已经确定细菌毒力因子的产生受到严格的生长阶段调节。现在很明显,细菌毒力的表达也受到在微环境中释放的真核信使的检测控制,这些信使作为内分泌或神经内分泌调节剂。在宿主生理应激的情况下,许多真核因子被释放并被革兰氏阴性菌检测到,而革兰氏阴性菌则迅速适应其生理机能。例如,铜绿假单胞菌可以结合宿主免疫系统的元素,如干扰素-γ和强啡肽,然后通过群体感应电路增强其毒力。大肠杆菌对儿茶酚胺家族神经激素的敏感性似乎通过最近发现的细菌肾上腺素受体来传递。在本综述中,我们将描述宿主产生的各种真核信号分子激活革兰氏阴性细菌毒力的机制。特别注意假单胞菌,其代表物种铜绿假单胞菌是一种常见的机会性病原体。讨论将特别侧重于这些新型病原体传感器从感应到涉及毒力因子调节的转导机制所发挥的关键作用。最后,我们将讨论宿主信号分子对与不同人体组织相关的共生或机会性病原体的影响。