Department of Palaeontology, Natural History Museum, London, United Kingdom.
PLoS One. 2012;7(5):e36904. doi: 10.1371/journal.pone.0036904. Epub 2012 May 22.
The most primitive ornithischian dinosaurs were small bipeds, but quadrupedality evolved three times independently in the clade. The transition to quadrupedality from bipedal ancestors is rare in the history of terrestrial vertebrate evolution, and extant analogues do not exist. Constraints imposed on quadrupedal ornithischians by their ancestral bipedal bauplan remain unexplored, and consequently, debate continues about their stance and gait. For example, it has been proposed that some ornithischians could run, while others consider that none were cursorial.
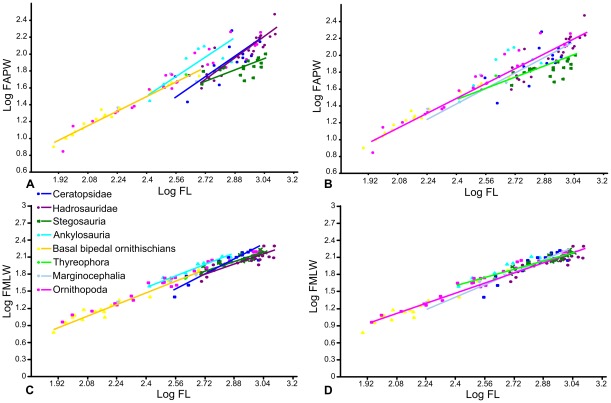
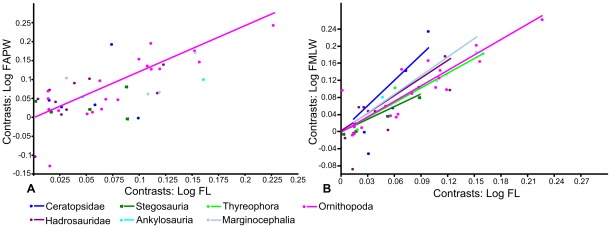
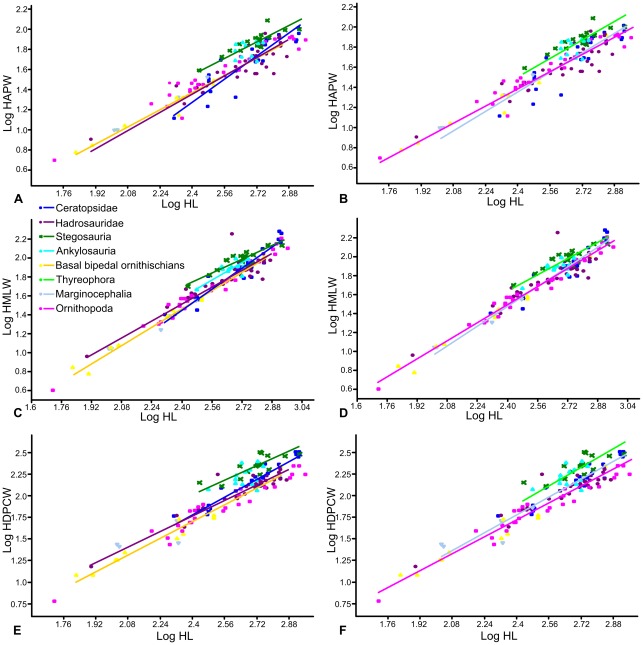
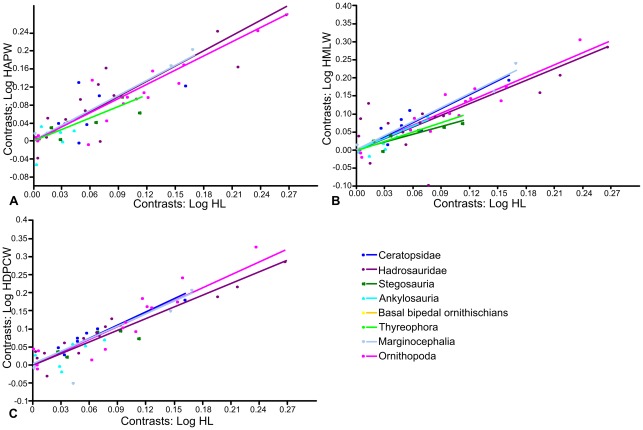
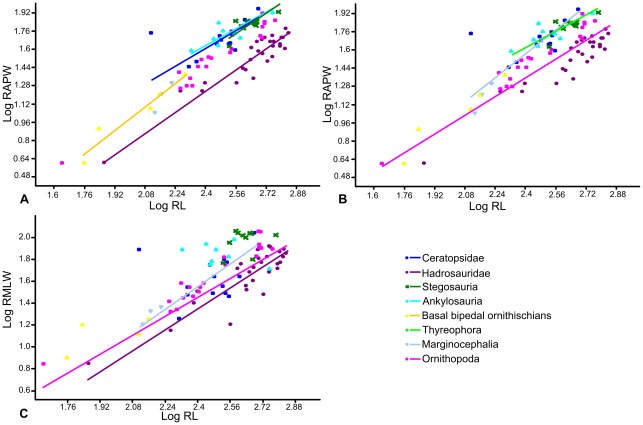
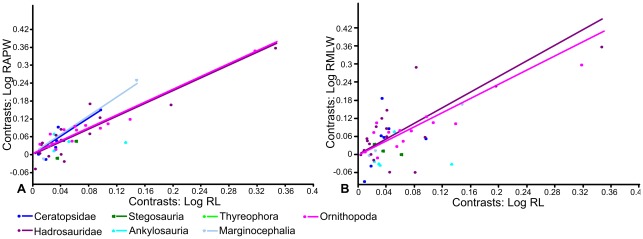
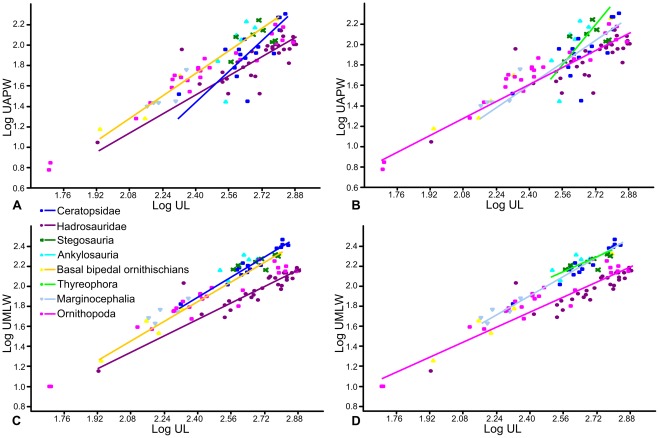
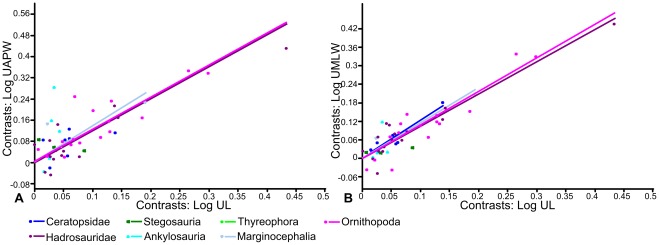
METHODOLOGY/PRINCIPAL FINDINGS: Drawing on biomechanical concepts of limb bone scaling and locomotor theory developed for extant taxa, we use the largest dataset of ornithischian postcranial measurements so far compiled to examine stance and gait in quadrupedal ornithischians. Differences in femoral midshaft eccentricity in hadrosaurs and ceratopsids may indicate that hadrosaurs placed their feet on the midline during locomotion, while ceratopsids placed their feet more laterally, under the hips. More robust humeri in the largest ceratopsids relative to smaller taxa may be due to positive allometry in skull size with body mass in ceratopsids, while slender humeri in the largest stegosaurs may be the result of differences in dermal armor distribution within the clade. Hadrosaurs are found to display the most cursorial morphologies of the quadrupedal ornithischian cades, indicating higher locomotor performance than in ceratopsids and thyreophorans.
CONCLUSIONS/SIGNIFICANCE: Limb bone scaling indicates that a previously unrealised diversity of stances and gaits were employed by quadrupedal ornithischians despite apparent convergence in limb morphology. Grouping quadrupedal ornithischians together as a single functional group hides this disparity. Differences in limb proportions and scaling are likely due to the possession of display structures such as horns, frills and dermal armor that may have affected the center of mass of the animal, and differences in locomotor behaviour such as migration, predator escape or home range size.
最原始的鸟脚亚目恐龙是小型两足动物,但该类群的四足性独立进化了三次。从两足祖先向四足过渡在陆地脊椎动物进化史上是罕见的,现存的类似物并不存在。其祖先两足动物形态所施加的对四足鸟脚亚目动物的限制仍未得到探索,因此,关于它们的姿势和步态的争论仍在继续。例如,有人提出一些鸟脚亚目恐龙可能会奔跑,而另一些人则认为没有一种是奔跑的。
方法/主要发现:借鉴为现生物种发展的肢骨比例和运动理论的生物力学概念,我们利用迄今为止最大的鸟脚亚目后肢测量数据集,研究了四足鸟脚亚目恐龙的姿势和步态。鸭嘴龙类和角龙类股骨中轴偏心度的差异可能表明,鸭嘴龙类在运动时将脚放在中线位置,而角龙类则将脚放在更靠外侧的位置,在臀部下方。最大的角龙类相对于较小的类群,肱骨更粗壮,这可能是由于角龙类头骨大小与体重呈正异速生长,而最大的剑龙类肱骨较细,这可能是由于该类群内真皮装甲分布的差异造成的。鸭嘴龙类被发现具有四足鸟脚亚目恐龙中最具奔跑形态的特征,表明其运动性能高于角龙类和基干鸟脚类。
结论/意义:肢骨比例表明,尽管四肢形态明显趋同,但四足鸟脚亚目恐龙采用了以前未被认识到的多种姿势和步态。将四足鸟脚亚目恐龙归为一个单一的功能群体掩盖了这种差异。四肢比例和比例的差异可能是由于拥有展示结构,如角、颈盾和真皮装甲,这些结构可能影响了动物的质心,以及运动行为的差异,如迁徙、逃避捕食者或栖息地范围的大小。