Jacqui Wood Cancer Centre, Division of Cancer Research, Ninewells Hospital and Medical School, University of Dundee, Dundee, Scotland, UK.
Oncogene. 2013 Aug 8;32(32):3765-81. doi: 10.1038/onc.2012.388. Epub 2012 Sep 10.
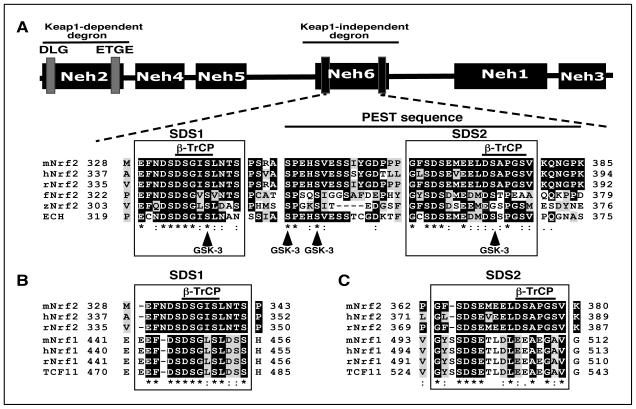
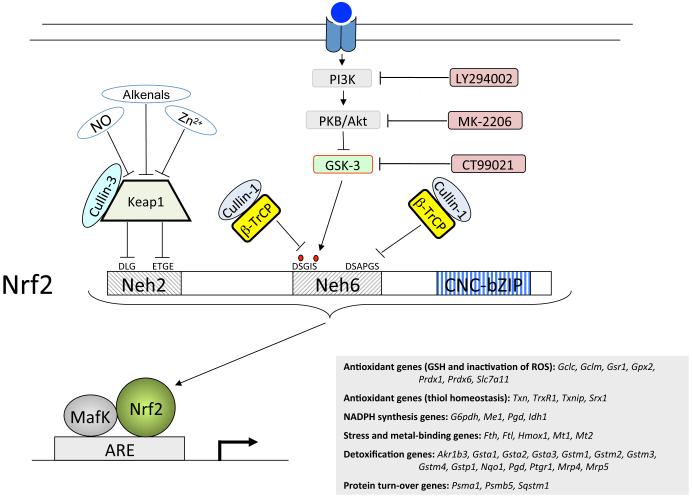
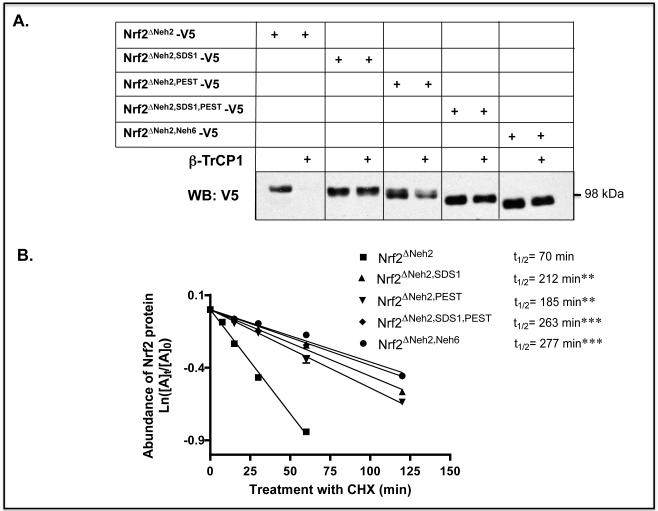
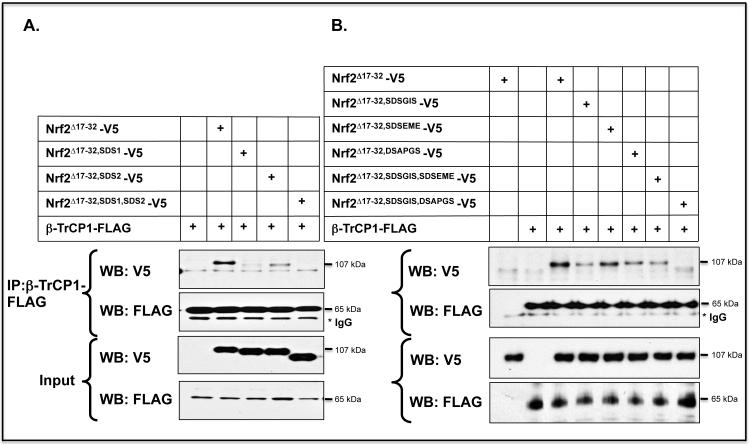
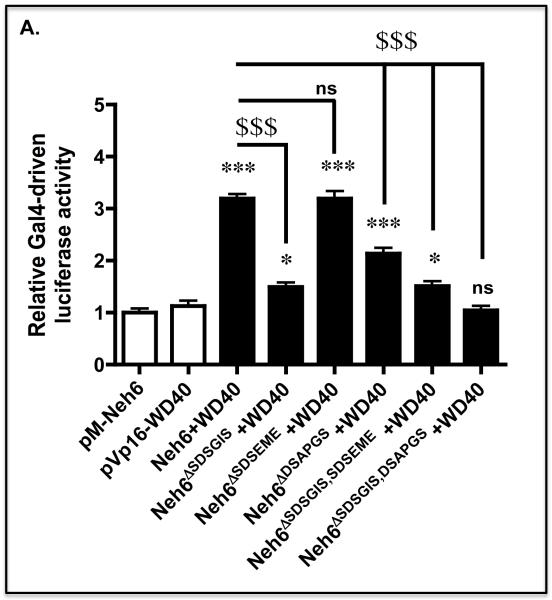
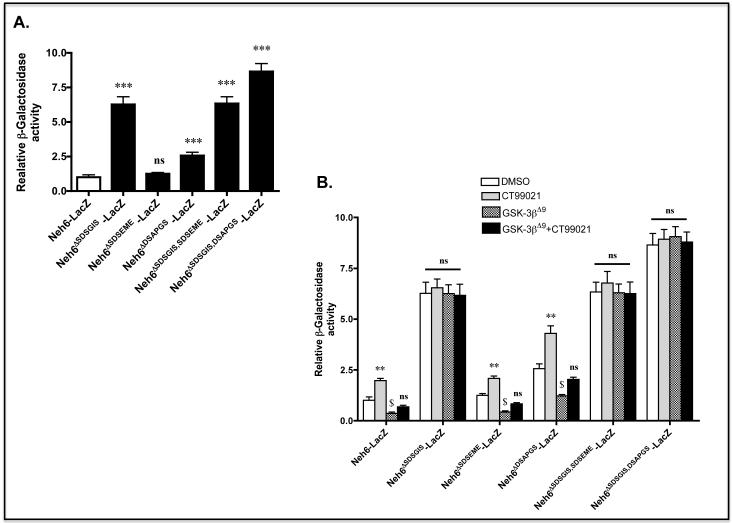
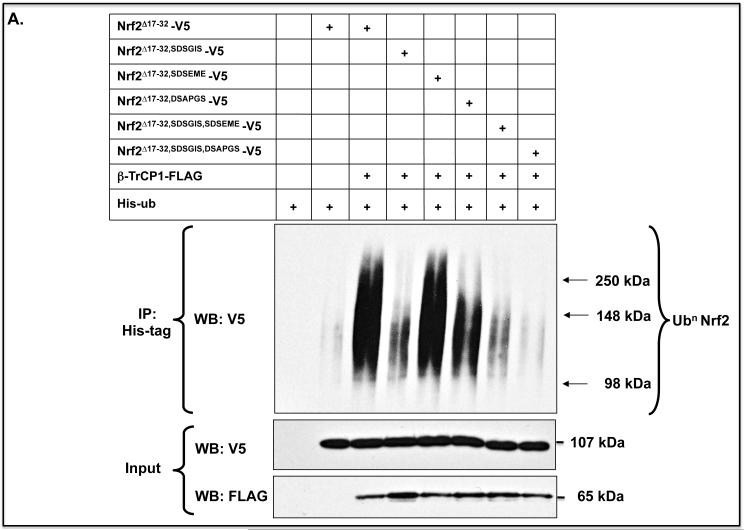
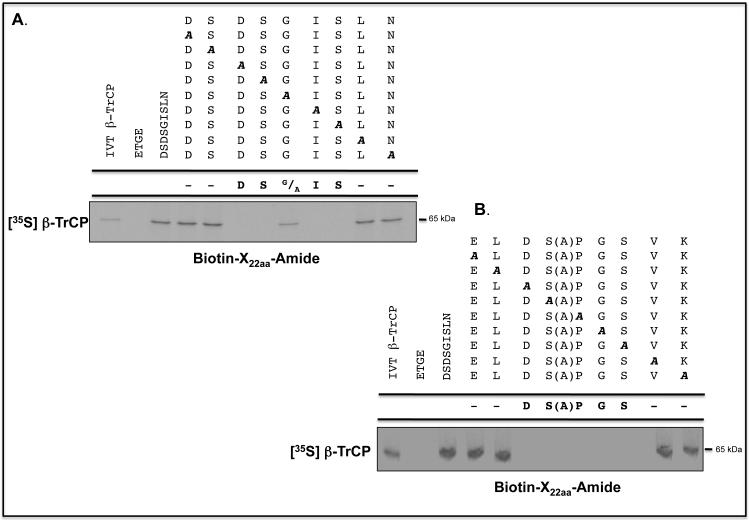
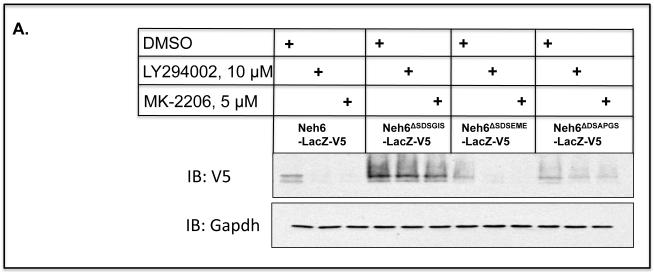
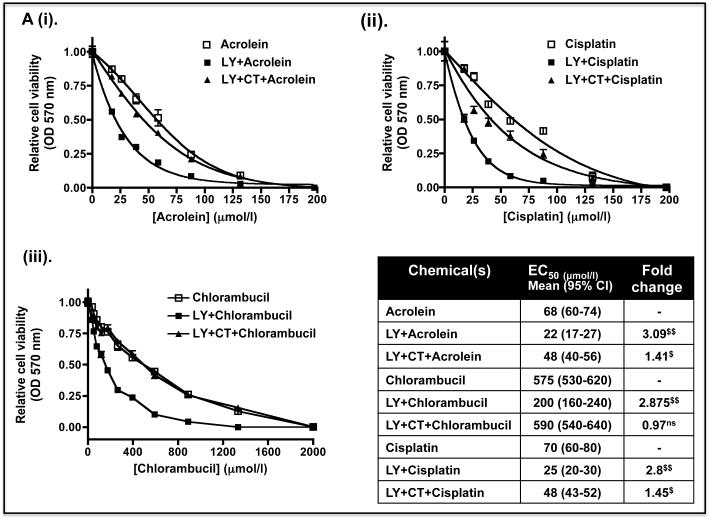
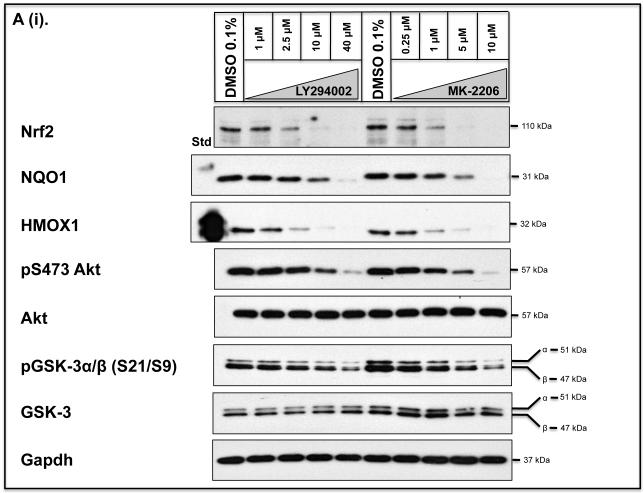
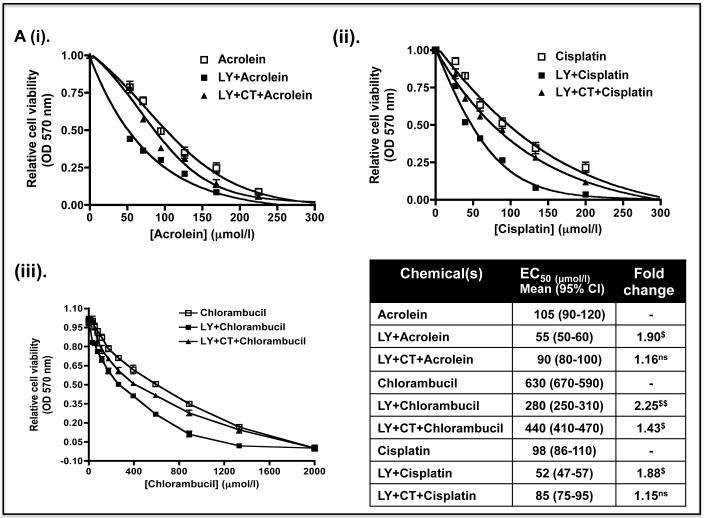
Identification of regulatable mechanisms by which transcription factor NF-E2 p45-related factor 2 (Nrf2) is repressed will allow strategies to be designed that counter drug resistance associated with its upregulation in tumours that harbour somatic mutations in Kelch-like ECH-associated protein-1 (Keap1), a gene that encodes a joint adaptor and substrate receptor for the Cul3-Rbx1/Roc1 ubiquitin ligase. We now show that mouse Nrf2 contains two binding sites for β-transducin repeat-containing protein (β-TrCP), which acts as a substrate receptor for the Skp1-Cul1-Rbx1/Roc1 ubiquitin ligase complex. Deletion of either binding site in Nrf2 decreased β-TrCP-mediated ubiquitylation of the transcription factor. The ability of one of the two β-TrCP-binding sites to serve as a degron could be both increased and decreased by manipulation of glycogen synthase kinase-3 (GSK-3) activity. Biotinylated-peptide pull-down assays identified DSGIS(338) and DSAPGS(378) as the two β-TrCP-binding motifs in Nrf2. Significantly, our pull-down assays indicated that β-TrCP binds a phosphorylated version of DSGIS more tightly than its non-phosphorylated counterpart, whereas this was not the case for DSAPGS. These data suggest that DSGIS, but not DSAPGS, contains a functional GSK-3 phosphorylation site. Activation of GSK-3 in Keap1-null mouse embryonic fibroblasts (MEFs), or in human lung A549 cells that contain mutant Keap1, by inhibition of the phosphoinositide 3-kinase (PI3K)-protein kinase B (PKB)/Akt pathway markedly reduced endogenous Nrf2 protein and decreased to 10-50% of normal the levels of mRNA for prototypic Nrf2-regulated enzymes, including the glutamate-cysteine ligase catalytic and modifier subunits, glutathione S-transferases Alpha-1 and Mu-1, haem oxygenase-1 and NAD(P)H:quinone oxidoreductase-1. Pre-treatment of Keap1(-/-) MEFs or A549 cells with the LY294002 PI3K inhibitor or the MK-2206 PKB/Akt inhibitor increased their sensitivity to acrolein, chlorambucil and cisplatin between 1.9-fold and 3.1-fold, and this was substantially attenuated by simultaneous pre-treatment with the GSK-3 inhibitor CT99021.
通过鉴定转录因子 NF-E2 p45 相关因子 2(Nrf2)受到抑制的调节机制,我们将能够设计出对抗策略,以应对与肿瘤中 Nrf2 上调相关的药物耐药性,这些肿瘤中存在 Kelch 样 ECH 相关蛋白 1(Keap1)的体细胞突变,该基因编码 Cul3-Rbx1/Roc1 泛素连接酶的联合接头和底物受体。我们现在表明,小鼠 Nrf2 含有两个结合β-转导素重复蛋白(β-TrCP)的结合位点,β-TrCP 作为 Skp1-Cul1-Rbx1/Roc1 泛素连接酶复合物的底物受体。Nrf2 中两个结合位点之一的缺失降低了转录因子的 β-TrCP 介导的泛素化。通过操纵糖原合成酶激酶-3(GSK-3)活性,可以增加和减少其中一个 β-TrCP 结合位点作为降解信号的能力。生物素化肽下拉测定鉴定出 DSGIS(338)和 DSAPGS(378)为 Nrf2 中的两个 β-TrCP 结合基序。重要的是,我们的下拉测定表明,β-TrCP 与磷酸化的 DSGIS 结合比其非磷酸化的对应物更紧密,而对于 DSAPGS 则不是这种情况。这些数据表明 DSGIS 而不是 DSAPGS 含有功能性 GSK-3 磷酸化位点。通过抑制磷酸肌醇 3-激酶(PI3K)-蛋白激酶 B(PKB)/ Akt 途径,在 Keap1 缺失的小鼠胚胎成纤维细胞(MEF)或含有突变 Keap1 的人肺 A549 细胞中激活 GSK-3,可显著降低内源性 Nrf2 蛋白,并将原代 Nrf2 调节酶的 mRNA 水平降低至正常的 10-50%,包括谷氨酸-半胱氨酸连接酶催化亚基和修饰亚基、谷胱甘肽 S-转移酶 Alpha-1 和 Mu-1、血红素加氧酶-1 和 NAD(P)H:醌氧化还原酶-1。用 LY294002 PI3K 抑制剂或 MK-2206 PKB/Akt 抑制剂预处理 Keap1(-/-)MEF 或 A549 细胞可将它们对丙烯醛、氯氨嘧啶和顺铂的敏感性增加 1.9 至 3.1 倍,并且同时用 GSK-3 抑制剂 CT99021 预处理可大大减弱这种敏感性。