Kaas Jon H
Department of Psychology, Vanderbilt University, Nashville, TN, USA.
Wiley Interdiscip Rev Cogn Sci. 2013 Jan;4(1):33-45. doi: 10.1002/wcs.1206. Epub 2012 Nov 8.
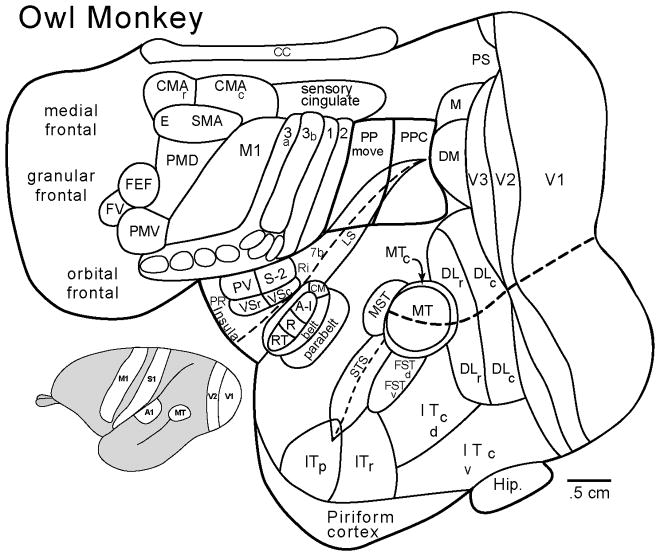
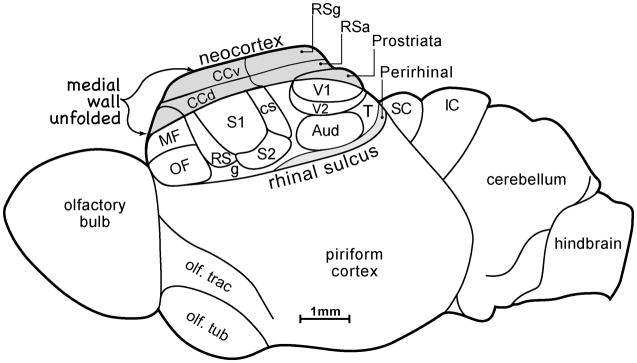
The large size and complex organization of the human brain makes it unique among primate brains. In particular, the neocortex constitutes about 80% of the brain, and this cortex is subdivided into a large number of functionally specialized regions, the cortical areas. Such a brain mediates accomplishments and abilities unmatched by any other species. How did such a brain evolve? Answers come from comparative studies of the brains of present-day mammals and other vertebrates in conjunction with information about brain sizes and shapes from the fossil record, studies of brain development, and principles derived from studies of scaling and optimal design. Early mammals were small, with small brains, an emphasis on olfaction, and little neocortex. Neocortex was transformed from the single layer of output pyramidal neurons of the dorsal cortex of earlier ancestors to the six layers of all present-day mammals. This small cap of neocortex was divided into 20-25 cortical areas, including primary and some of the secondary sensory areas that characterize neocortex in nearly all mammals today. Early placental mammals had a corpus callosum connecting the neocortex of the two hemispheres, a primary motor area, M1, and perhaps one or more premotor areas. One line of evolution, Euarchontoglires, led to present-day primates, tree shrews, flying lemurs, rodents, and rabbits. Early primates evolved from small-brained, nocturnal, insect-eating mammals with an expanded region of temporal visual cortex. These early nocturnal primates were adapted to the fine branch niche of the tropical rainforest by having an even more expanded visual system that mediated visually guided reaching and grasping of insects, small vertebrates, and fruits. Neocortex was greatly expanded and included an array of cortical areas that characterize neocortex of all living primates. Specializations of the visual system included new visual areas that contributed to a dorsal stream of visuomotor processing in a greatly enlarged region of posterior parietal cortex and an expanded motor system and the addition of a ventral premotor area. Higher visual areas in a large temporal lobe facilitated object recognition, and frontal cortex included granular prefrontal cortex. Auditory cortex included the primary and secondary auditory areas that characterize prosimian and anthropoid primates today. As anthropoids emerged as diurnal primates, the visual system specialized for detailed foveal vision. Other adaptations included an expansion of prefrontal cortex and insular cortex. The human and chimpanzee-bonobo lineages diverged some 6-8 million years ago with brains that were about one third the size of modern humans. Over the last 2 million years, the brains of our more recent ancestors increased greatly in size, especially in the prefrontal, posterior parietal, lateral temporal, and insular regions. Specialization of the two cerebral hemispheres for related, but different functions became pronounced, and language and other impressive cognitive abilities emerged. WIREs Cogn Sci 2013, 4:33-45. doi: 10.1002/wcs.1206 This article is categorized under: Neuroscience > Anatomy.
人类大脑的庞大尺寸和复杂结构使其在灵长类动物的大脑中独树一帜。特别是,新皮层约占大脑的80%,且该皮层又被细分为大量功能特化的区域,即皮质区。这样的大脑介导了其他任何物种都无法比拟的成就和能力。这样的大脑是如何进化的呢?答案来自对当今哺乳动物和其他脊椎动物大脑的比较研究,结合化石记录中有关大脑大小和形状的信息、大脑发育研究,以及从缩放和最优设计研究中得出的原理。早期哺乳动物体型较小,大脑也小,侧重于嗅觉,新皮层很少。新皮层从早期祖先背侧皮层的单层输出锥体神经元转变为所有现代哺乳动物的六层结构。这块小小的新皮层帽被划分为20 - 25个皮质区,包括几乎所有当今哺乳动物新皮层所特有的初级和一些次级感觉区。早期胎盘哺乳动物有胼胝体连接两个半球的新皮层、一个初级运动区M1,或许还有一个或多个运动前区。一条进化路线,即真灵长大目,演化出了当今的灵长类动物、树鼩、鼯猴、啮齿动物和兔子。早期灵长类动物由大脑较小、夜行性、食虫的哺乳动物进化而来,其颞部视觉皮层区域有所扩大。这些早期夜行性灵长类动物通过拥有一个更加扩展的视觉系统适应了热带雨林的细枝生态位,该视觉系统介导视觉引导下对昆虫、小型脊椎动物和果实的抓取。新皮层大幅扩展,包括一系列所有现存灵长类动物新皮层所特有的皮质区。视觉系统的特化包括新的视觉区,这些区域在大大扩大的后顶叶皮层区域促成了视觉运动处理的背侧流,以及一个扩展的运动系统和一个腹侧运动前区的增加。大颞叶中的高级视觉区有助于物体识别,额叶皮层包括颗粒状前额叶皮层。听觉皮层包括当今原猴亚目和类人猿灵长类动物所特有的初级和次级听觉区。随着类人猿成为昼行性灵长类动物,视觉系统专门用于精细的中央凹视觉。其他适应性变化包括前额叶皮层和岛叶皮层的扩展。人类和黑猩猩 - 倭黑猩猩谱系在约600 - 800万年前分化,当时的大脑大小约为现代人类的三分之一。在过去的200万年里,我们近代祖先的大脑尺寸大幅增加,尤其是在前额叶、后顶叶、外侧颞叶和岛叶区域。两个大脑半球在相关但不同功能上的特化变得显著,语言和其他令人印象深刻的认知能力也随之出现。WIREs认知科学2013年,4:33 - 45。doi:10.1002/wcs.1206 本文分类如下:神经科学>解剖学