Department of Cell and Developmental Biology, John Innes Centre, Norwich Research Park, Norwich, United Kingdom.
PLoS Biol. 2013;11(4):e1001550. doi: 10.1371/journal.pbio.1001550. Epub 2013 Apr 30.
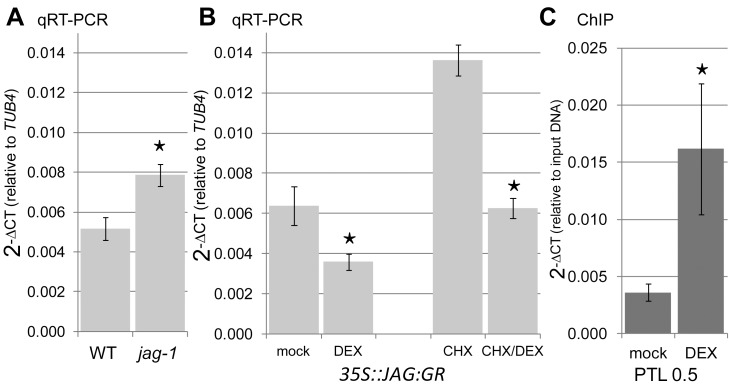
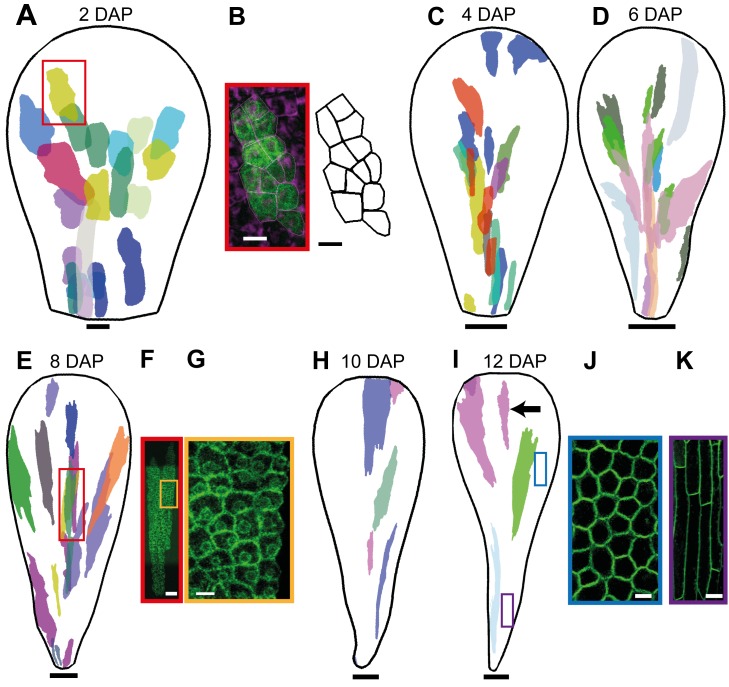
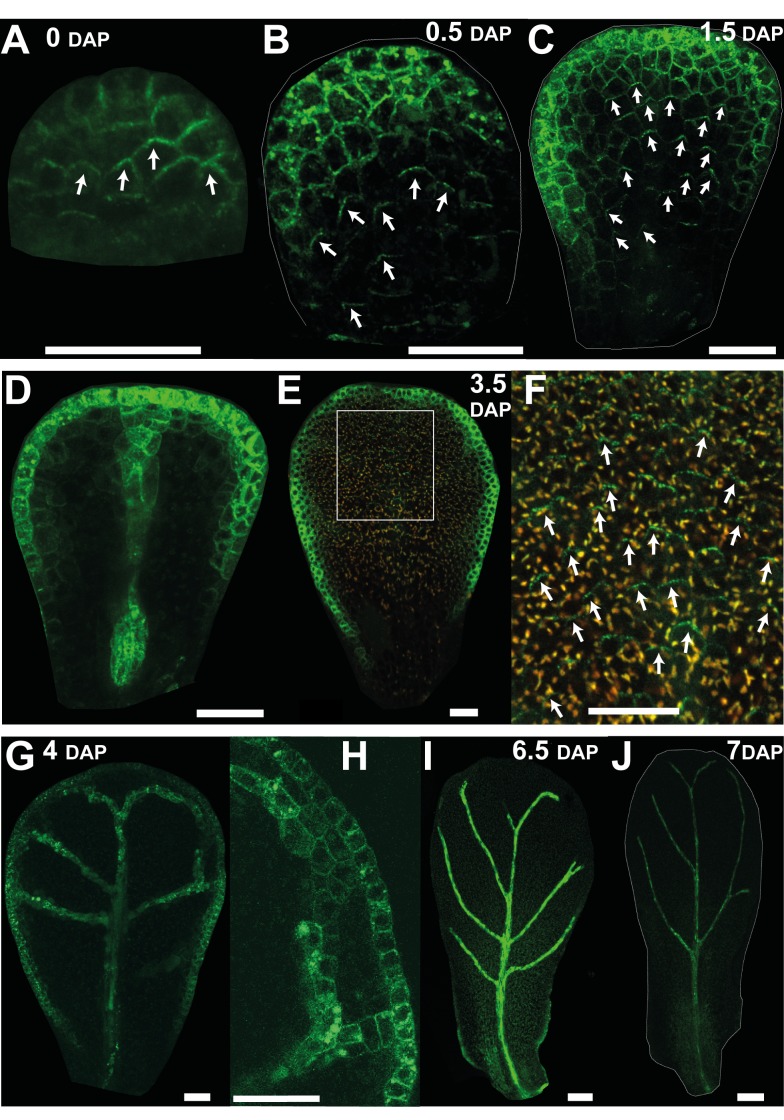
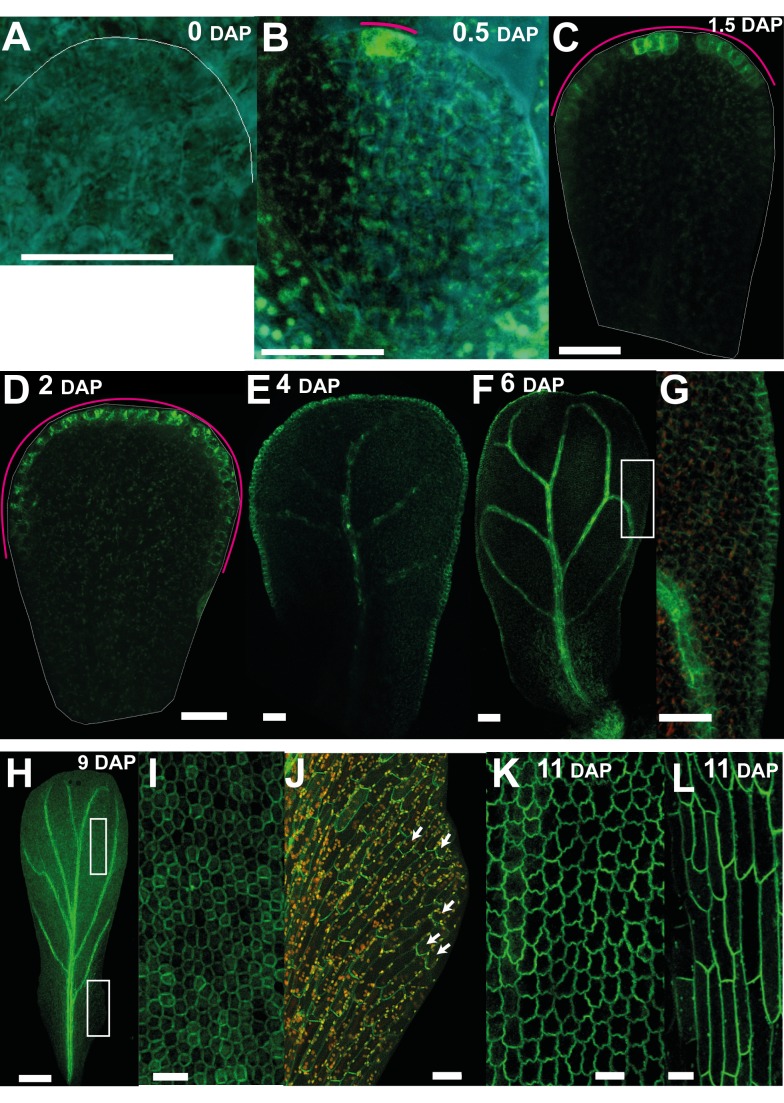
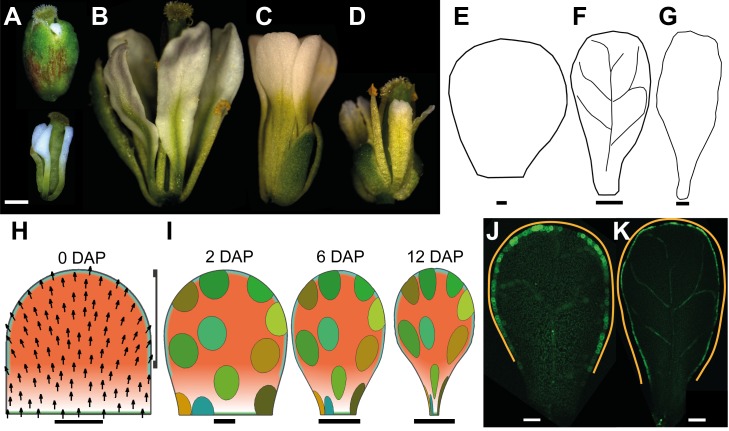
A flowering plant generates many different organs such as leaves, petals, and stamens, each with a particular function and shape. These types of organ are thought to represent variations on a common underlying developmental program. However, it is unclear how this program is modulated under different selective constraints to generate the diversity of forms observed. Here we address this problem by analysing the development of Arabidopsis petals and comparing the results to models of leaf development. We show that petal development involves a divergent polarity field with growth rates perpendicular to local polarity increasing towards the distal end of the petal. The hypothesis is supported by the observed pattern of clones induced at various stages of development and by analysis of polarity markers, which show a divergent pattern. We also show that JAGGED (JAG) has a key role in promoting distal enhancement of growth rates and influences the extent of the divergent polarity field. Furthermore, we reveal links between the polarity field and auxin function: auxin-responsive markers such as DR5 have a broader distribution along the distal petal margin, consistent with the broad distal organiser of polarity, and PETAL LOSS (PTL), which has been implicated in the control of auxin dynamics during petal initiation, is directly repressed by JAG. By comparing these results with those from studies on leaf development, we show how simple modifications of an underlying developmental system may generate distinct forms, providing flexibility for the evolution of different organ functions.
开花植物会生成许多不同的器官,如叶子、花瓣和雄蕊,每个器官都具有特定的功能和形状。这些类型的器官被认为代表了同一基本发育程序的不同变体。然而,目前尚不清楚该程序如何在不同的选择压力下进行调节,以产生所观察到的多样化形式。在这里,我们通过分析拟南芥花瓣的发育并将结果与叶片发育模型进行比较来解决这个问题。我们表明,花瓣的发育涉及一个发散的极性场,其生长速率垂直于局部极性,朝着花瓣的远端增加。这一假说得到了在不同发育阶段诱导的克隆模式和极性标记分析的支持,这些分析显示出发散的模式。我们还表明,JAGGED(JAG)在促进生长速率的远端增强方面起着关键作用,并影响发散极性场的范围。此外,我们揭示了极性场和生长素功能之间的联系:生长素反应标记物,如 DR5,在整个花瓣远端边缘的分布更广泛,与广泛的远端极性组织者一致,而在花瓣起始过程中控制生长素动力学的 PETAL LOSS(PTL)则直接受到 JAG 的抑制。通过将这些结果与叶片发育研究的结果进行比较,我们展示了如何通过对基本发育系统进行简单修改,从而产生不同的形式,为不同器官功能的进化提供了灵活性。