Greenebaum Cancer Center, University of Maryland, Baltimore, Maryland, United States of America.
PLoS One. 2013 May 13;8(5):e62826. doi: 10.1371/journal.pone.0062826. Print 2013.
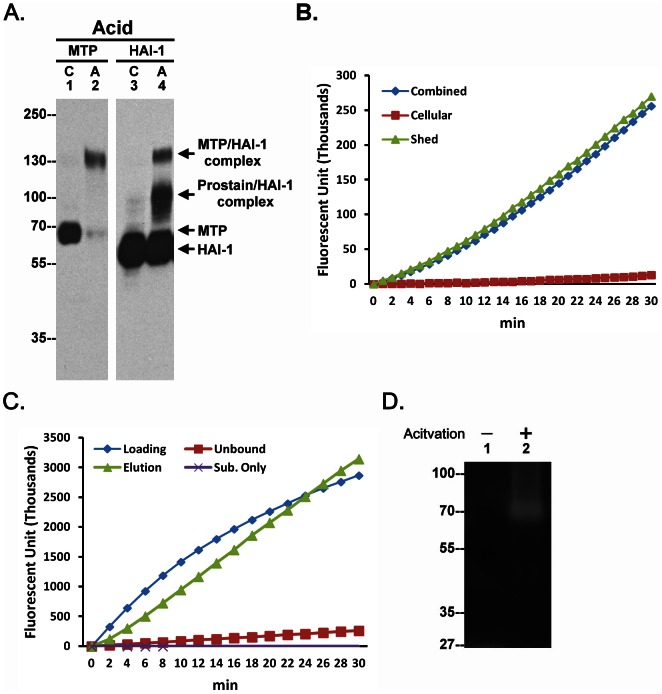
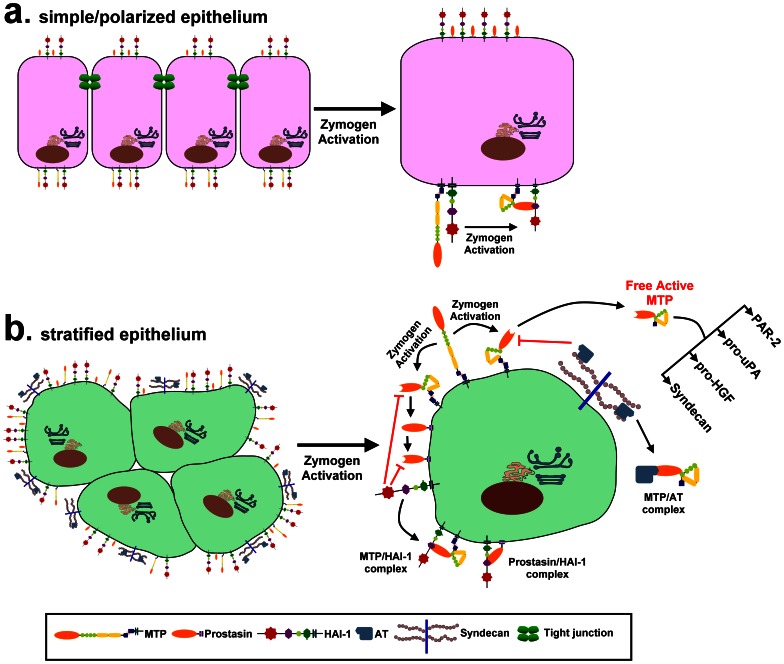
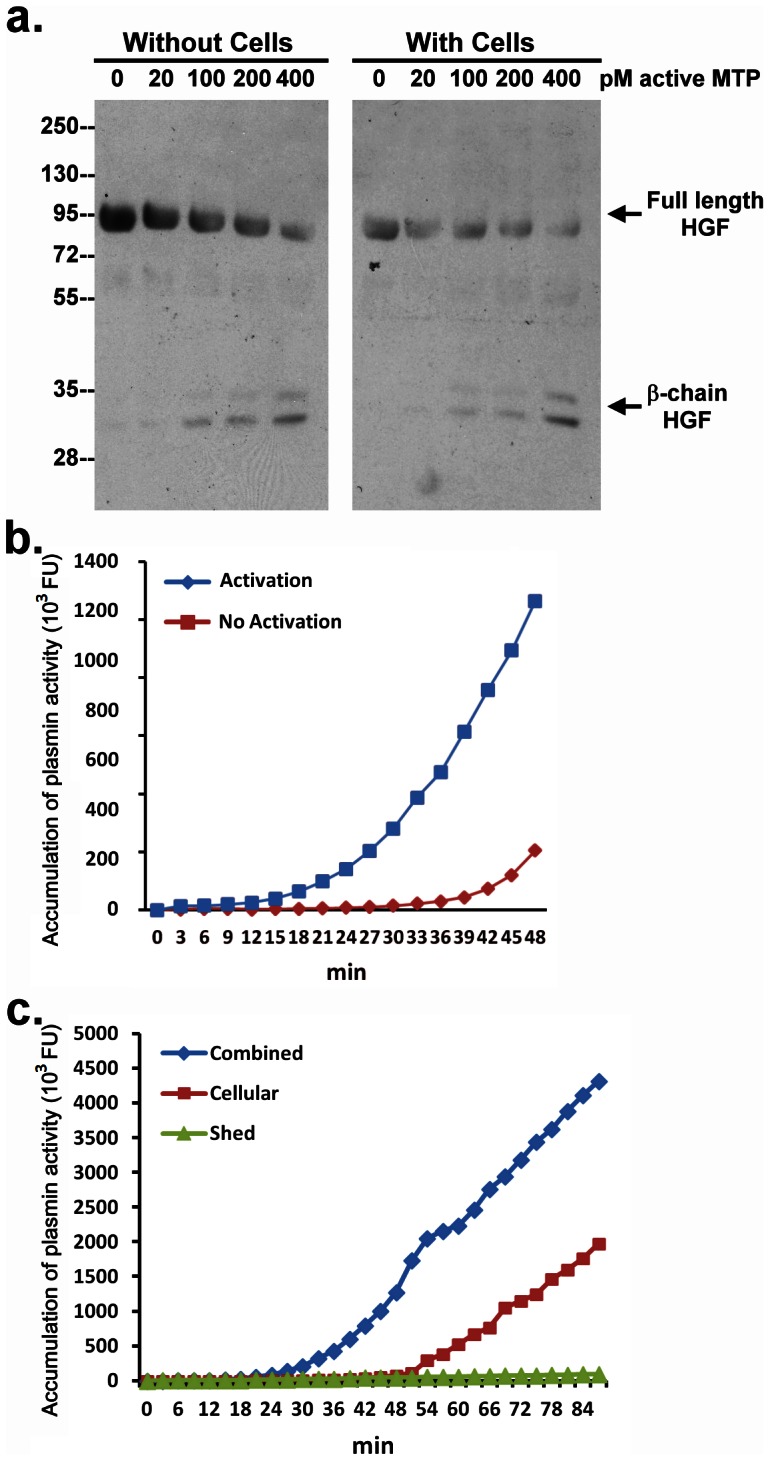
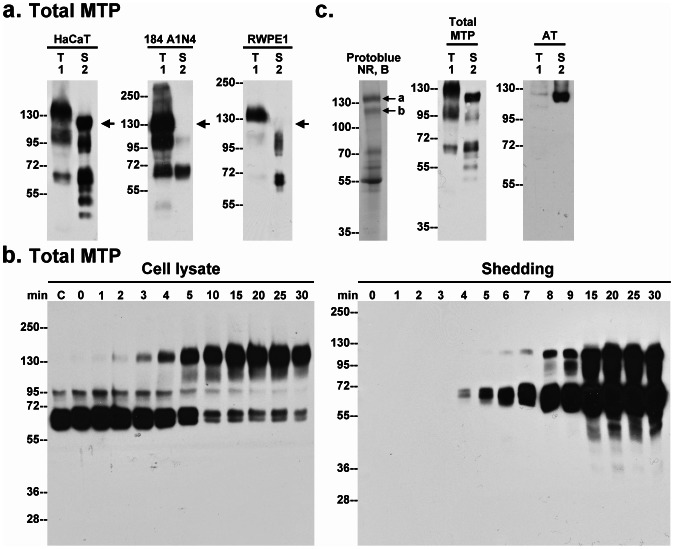
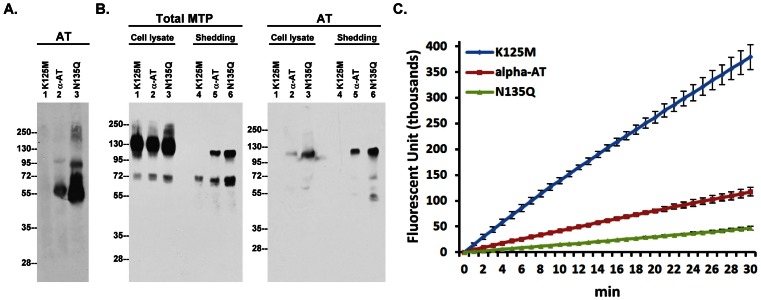
Matriptase, a membrane-associated serine protease, plays an essential role in epidermal barrier function through activation of the glycosylphosphatidylinositol (GPI)-anchored serine protease prostasin. The matriptase-prostasin proteolytic cascade is tightly regulated by hepatocyte growth factor activator inhibitor (HAI)-1 such that matriptase autoactivation and prostasin activation occur simultaneously and are followed immediately by the inhibition of both enzymes by HAI-1. However, the mechanisms whereby matriptase acts on extracellular substrates remain elusive. Here we report that some active matriptase can escape HAI-1 inhibition by being rapidly shed from the cell surface. In the pericellular environment, shed active matriptase is able to activate hepatocyte growth factor (HGF), accelerate plasminogen activation, and shed syndecan 1. The amount of active matriptase shed is inversely correlated with the amount of antithrombin (AT) bound to the surface of the keratinocytes. Binding of AT to the surface of keratinocytes is dependent on a functional heparin binding site, Lys-125, and that the N-glycosylation site Asn-135 be unglycosylated. This suggests that β-AT, and not α-AT, is responsible for regulation of pericellular matriptase activity in keratinocytes. Keratinocytes appear to rely on AT to regulate the level of pericellular active matriptase much more than breast and prostate epithelial cells in which AT regulation of matriptase activity occurs at much lower levels than keratinocytes. These results suggest that keratinocytes employ two distinct serine protease inhibitors to control the activation and processing of two different sets of matriptase substrates leading to different biological events: 1) HAI-1 for prostasin activation/inhibition, and 2) AT for the pericellular proteolysis involved in HGF activation, accelerating plasminogen activation, and shedding of syndecans.
组织蛋白酶 G 是一种膜相关丝氨酸蛋白酶,通过激活糖基磷脂酰肌醇 (GPI) 锚定丝氨酸蛋白酶原蛋白酶,在表皮屏障功能中发挥重要作用。组织蛋白酶 G-原蛋白酶蛋白酶级联反应受到肝细胞生长因子激活抑制剂 (HAI)-1 的严密调节,使得组织蛋白酶 G 自动激活和原蛋白酶激活同时发生,并立即被 HAI-1 抑制两种酶。然而,组织蛋白酶 G 作用于细胞外底物的机制仍然难以捉摸。在这里,我们报告说,一些活性组织蛋白酶 G 可以通过从细胞表面迅速脱落而逃脱 HAI-1 的抑制。在细胞外环境中,脱落的活性组织蛋白酶 G 能够激活肝细胞生长因子 (HGF),加速纤溶酶原激活,并脱落 syndecan 1。脱落的活性组织蛋白酶 G 的量与结合到角蛋白细胞表面的抗凝血酶 (AT) 的量成反比。AT 与角蛋白细胞表面的结合依赖于功能肝素结合位点 Lys-125,并且 N-糖基化位点 Asn-135 未糖基化。这表明β-AT,而不是α-AT,对角质形成细胞中细胞外组织蛋白酶 G 活性的调节负责。角质形成细胞似乎依赖 AT 来调节细胞外活性组织蛋白酶 G 的水平,远远超过乳腺和前列腺上皮细胞,在这些细胞中,AT 对组织蛋白酶 G 活性的调节发生在比角质形成细胞低得多的水平。这些结果表明,角质形成细胞采用两种不同的丝氨酸蛋白酶抑制剂来控制两组不同的组织蛋白酶 G 底物的激活和加工,从而导致不同的生物学事件:1)HAI-1 用于原蛋白酶激活/抑制,2)AT 用于涉及 HGF 激活、加速纤溶酶原激活和 syndecan 脱落的细胞外蛋白水解。