Zoological Museum, The Natural History Museum of Denmark, University of Copenhagen, Universitetsparken 15, DK-2100, Copenhagen, Denmark.
BMC Evol Biol. 2013 Aug 16;13:171. doi: 10.1186/1471-2148-13-171.
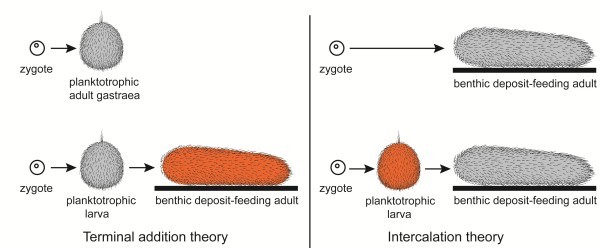
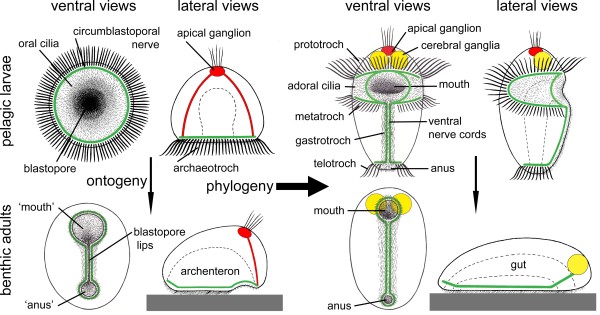
Two theories for the origin of animal life cycles with planktotrophic larvae are now discussed seriously: The terminal addition theory proposes a holopelagic, planktotrophic gastraea as the ancestor of the eumetazoans with addition of benthic adult stages and retention of the planktotrophic stages as larvae, i.e. the ancestral life cycles were indirect. The intercalation theory now proposes a benthic, deposit-feeding gastraea as the bilaterian ancestor with a direct development, and with planktotrophic larvae evolving independently in numerous lineages through specializations of juveniles.
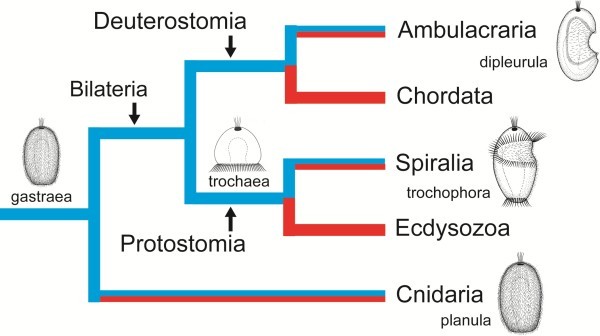
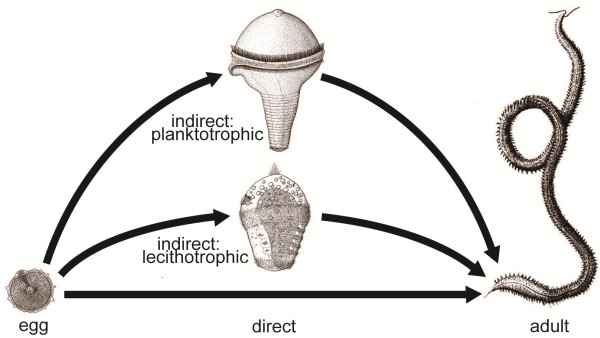
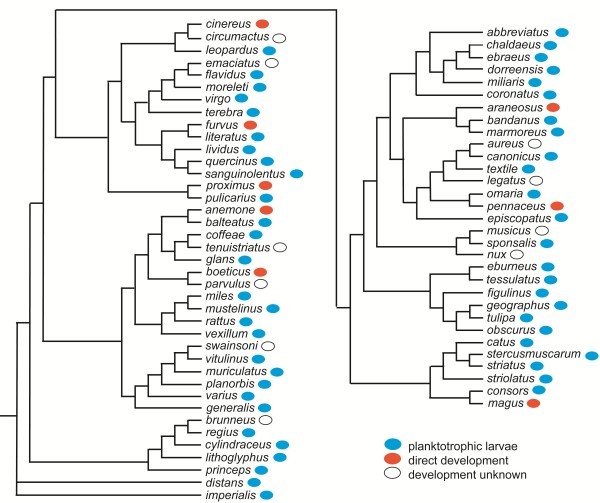
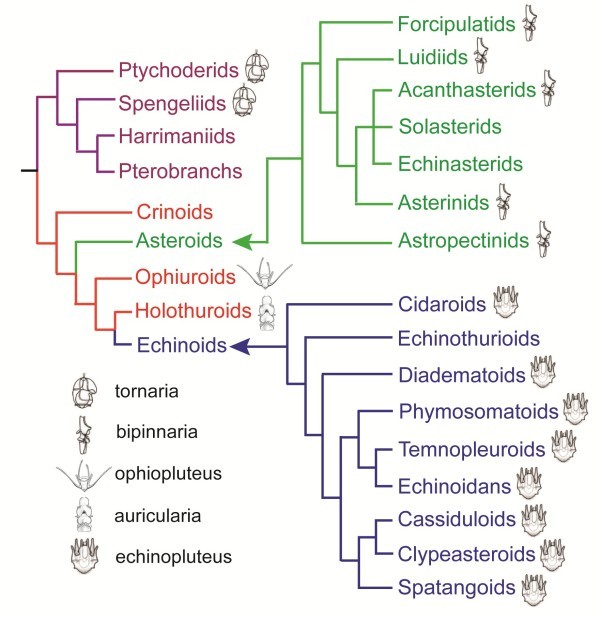
Information from the fossil record, from mapping of developmental types onto known phylogenies, from occurrence of apical organs, and from genetics gives no direct information about the ancestral eumetazoan life cycle; however, there are plenty of examples of evolution from an indirect development to direct development, and no unequivocal example of evolution in the opposite direction. Analyses of scenarios for the two types of evolution are highly informative. The evolution of the indirect spiralian life cycle with a trochophora larva from a planktotrophic gastraea is explained by the trochophora theory as a continuous series of ancestors, where each evolutionary step had an adaptational advantage. The loss of ciliated larvae in the ecdysozoans is associated with the loss of outer ciliated epithelia. A scenario for the intercalation theory shows the origin of the planktotrophic larvae of the spiralians through a series of specializations of the general ciliation of the juvenile. The early steps associated with the enhancement of swimming seem probable, but the following steps which should lead to the complicated downstream-collecting ciliary system are without any advantage, or even seem disadvantageous, until the whole structure is functional. None of the theories account for the origin of the ancestral deuterostome (ambulacrarian) life cycle.
All the available information is strongly in favor of multiple evolution of non-planktotrophic development, and only the terminal addition theory is in accordance with the Darwinian theory by explaining the evolution through continuous series of adaptational changes. This implies that the ancestor of the eumetazoans was a holopelagic, planktotrophic gastraea, and that the adult stages of cnidarians (sessile) and bilaterians (creeping) were later additions to the life cycle. It further implies that the various larval types are of considerable phylogenetic value.
目前有两种理论可以解释动物生命周期和浮游幼虫的起源:末端添加理论提出,一个全海洋浮游、浮游幼虫的后生动物作为有添加底栖成体阶段和保留浮游幼虫阶段的后生动物祖先,即祖先的生命周期是间接的。插入理论现在提出,一个底栖、沉积食性的原肠胚作为两侧对称动物的祖先,具有直接发育,并且通过幼体的特化,浮游幼虫在许多谱系中独立进化。
来自化石记录、发育类型映射到已知系统发育、顶端器官的出现以及遗传学的信息并没有直接提供关于祖先后生动物生命周期的信息;然而,有很多从间接发育到直接发育的进化例子,没有明确的相反方向进化的例子。对这两种进化类型的情景分析非常有启发性。从浮游幼虫的螺旋形间接生命循环的进化,通过原肠胚理论解释为一个连续的祖先系列,其中每一个进化步骤都有适应优势。蜕皮动物中纤毛幼虫的丧失与外纤毛上皮的丧失有关。插入理论的一个情景表明,螺旋形动物的浮游幼虫起源于幼体一般纤毛的一系列特化。与增强游泳能力相关的早期步骤似乎是可能的,但是随后的步骤应该导致复杂的下游收集纤毛系统,没有任何优势,甚至似乎是不利的,直到整个结构是功能的。这两种理论都没有解释祖先后口动物(步带动物)生命周期的起源。
所有可用的信息都强烈支持非浮游幼虫发育的多向进化,只有末端添加理论通过解释通过连续的适应变化的进化与达尔文理论相符。这意味着后生动物的祖先是一个全海洋浮游、浮游幼虫的后生动物,刺胞动物(固着)和两侧对称动物(爬行)的成体阶段是生命周期的后来添加。这进一步意味着各种幼虫类型具有相当大的系统发育价值。