Department of Chemistry, Centre for Chemical Biology, Krebs Institute, University of Sheffield, Sheffield, S3 7HF, UK and EaStCHEM School of Chemistry and Collaborative Optical Spectroscopy, Micromanipulation and Imaging Centre, The University of Edinburgh, West Mains Road, Edinburgh EH9 3JJ, UK.
Nucleic Acids Res. 2013 Nov;41(21):9839-47. doi: 10.1093/nar/gkt737. Epub 2013 Aug 23.
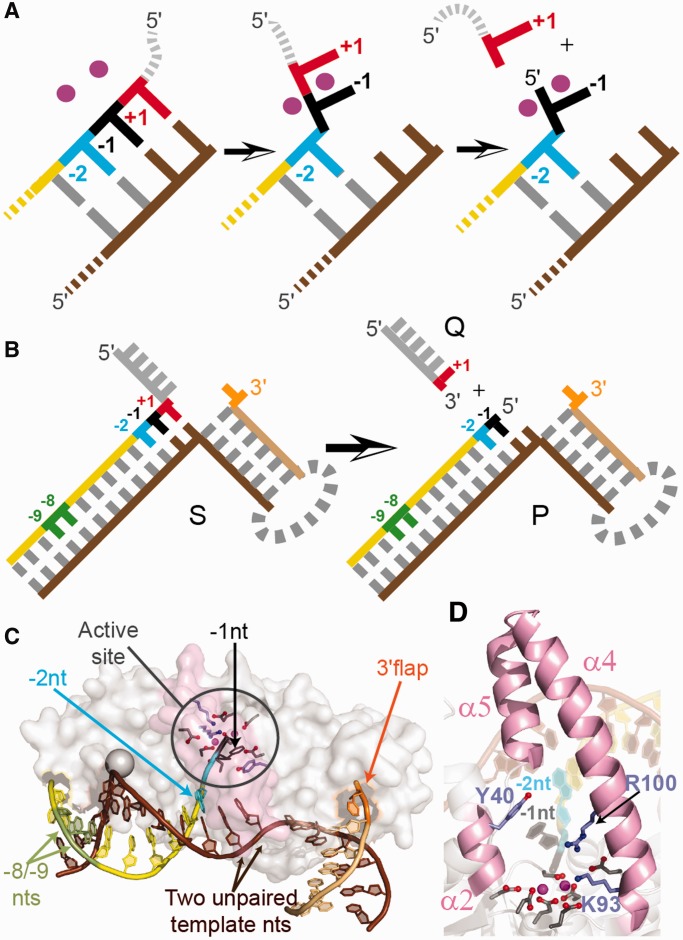
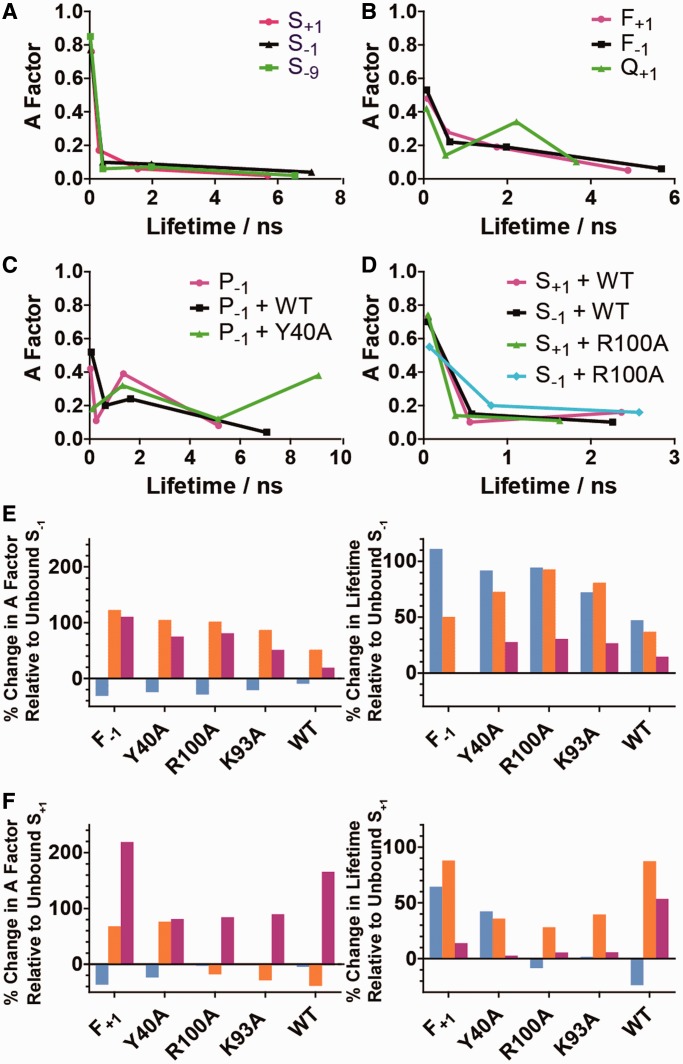
The structure- and strand-specific phosphodiesterase flap endonuclease-1 (FEN1), the prototypical 5'-nuclease, catalyzes the essential removal of 5'-single-stranded flaps during replication and repair. FEN1 achieves this by selectively catalyzing hydrolysis one nucleotide into the duplex region of substrates, always targeting the 5'-strand. This specificity is proposed to arise by unpairing the 5'-end of duplex to permit the scissile phosphate diester to contact catalytic divalent metal ions. Providing the first direct evidence for this, we detected changes induced by human FEN1 (hFEN1) in the low-energy CD spectra and fluorescence lifetimes of 2-aminopurine in substrates and products that were indicative of unpairing. Divalent metal ions were essential for unpairing. However, although 5'-nuclease superfamily-conserved active-site residues K93 and R100 were required to produce unpaired product, they were not necessary to unpair substrates. Nevertheless, a unique arrangement of protein residues around the unpaired DNA was detected only with wild-type protein, suggesting a cooperative assembly of active-site residues that may be triggered by unpaired DNA. The general principles of FEN1 strand and reaction-site selection, which depend on the ability of juxtaposed divalent metal ions to unpair the end of duplex DNA, may also apply more widely to other structure- and strand-specific nucleases.
结构和链特异性磷酸二酯酶 flap 内切核酸酶 1(FEN1),即典型的 5'-核酸酶,在复制和修复过程中催化必需的去除 5'-单链 flap。FEN1 通过选择性地催化水解一个核苷酸进入底物的双链区域来实现这一点,始终针对 5'-链。这种特异性被认为是通过使双链的 5'-末端解旋来允许切割磷酸二酯键与催化的二价金属离子接触而产生的。我们首次直接证明了这一点,我们检测到了人 FEN1(hFEN1)在底物和产物中 2-氨基嘌呤的低能量 CD 光谱和荧光寿命的变化,这些变化表明存在解旋。二价金属离子对于解旋是必需的。然而,尽管 5'-核酸酶超家族保守的活性位点残基 K93 和 R100 需要产生未配对的产物,但它们对于解旋底物不是必需的。尽管如此,只有野生型蛋白才能检测到围绕未配对 DNA 的独特蛋白残基排列,这表明活性位点残基的协同组装可能是由未配对 DNA 触发的。FEN1 链和反应位点选择的一般原则取决于相邻二价金属离子解开双链 DNA 末端的能力,这些原则也可能更广泛地适用于其他结构和链特异性核酸酶。