Burroughs Alexander Maxwell, Ando Yoshinari, Aravind L
National Center for Biotechnology Information, National Library of Medicine, National Institutes of Health, Bethesda, MD, USA.
Wiley Interdiscip Rev RNA. 2014 Mar-Apr;5(2):141-81. doi: 10.1002/wrna.1210. Epub 2013 Dec 5.
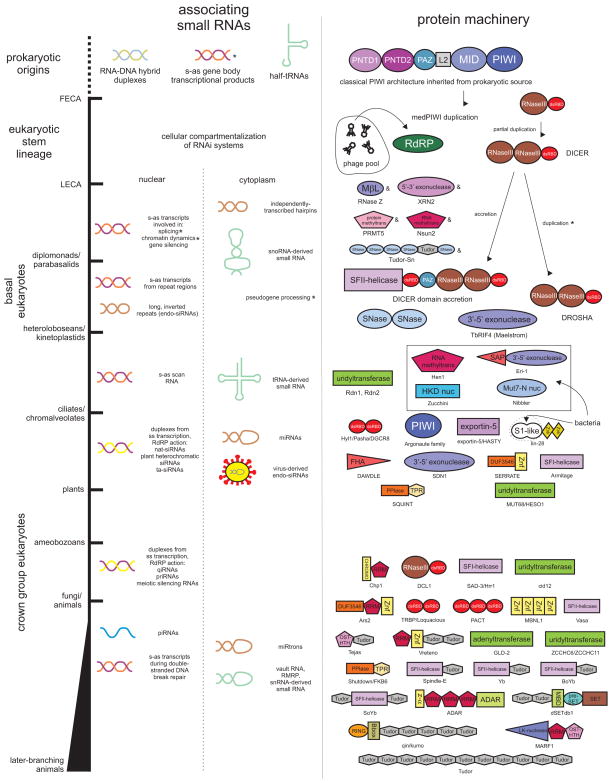
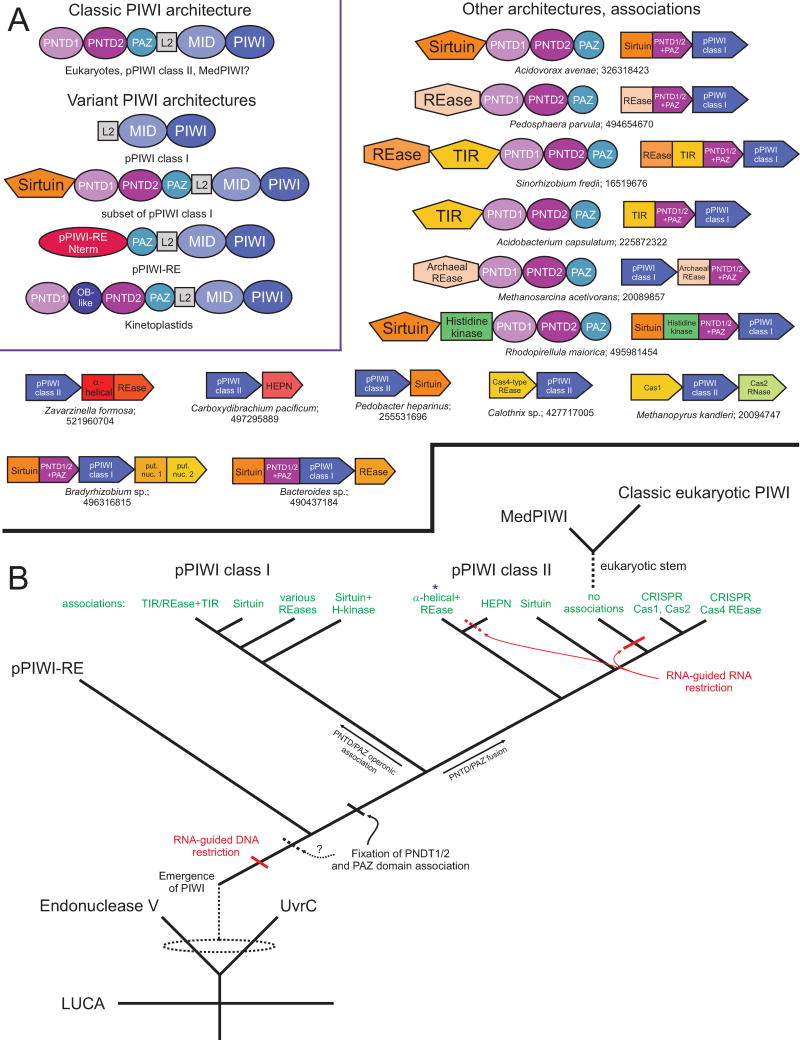
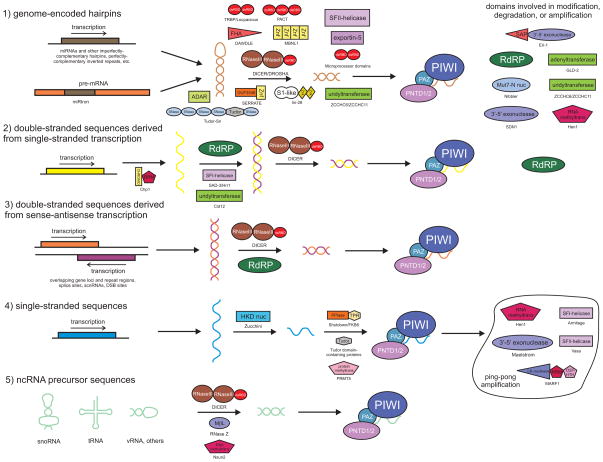
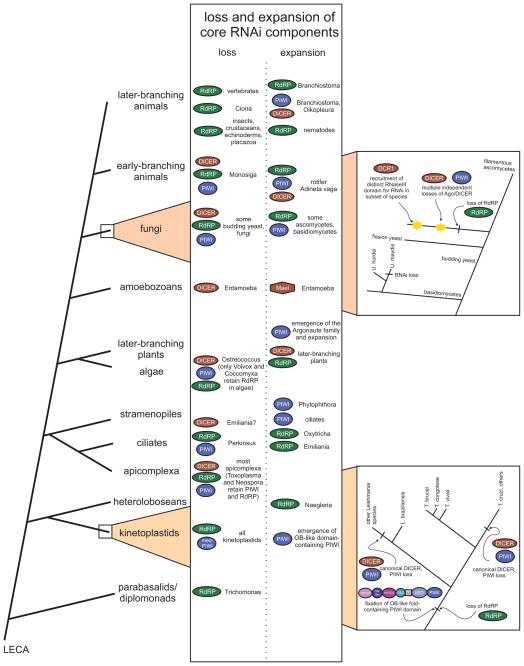
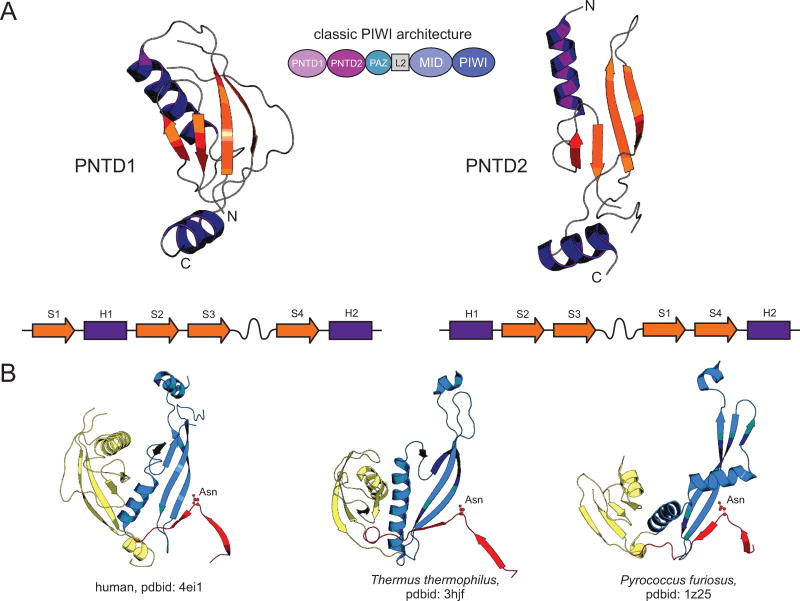
Our understanding of the pervasive involvement of small RNAs in regulating diverse biological processes has been greatly augmented by recent application of deep-sequencing technologies to small RNA across diverse eukaryotes. We review the currently known small RNA classes and place them in context of the reconstructed evolutionary history of the RNA interference (RNAi) protein machinery. This synthesis indicates that the earliest versions of eukaryotic RNAi systems likely utilized small RNA processed from three types of precursors: (1) sense-antisense transcriptional products, (2) genome-encoded, imperfectly complementary hairpin sequences, and (3) larger noncoding RNA precursor sequences. Structural dissection of PIWI proteins along with recent discovery of novel families (including Med13 of the Mediator complex) suggest that emergence of a distinct architecture with the N-terminal domains (also occurring separately fused to endoDNases in prokaryotes) formed via duplication of an ancestral unit was key to their recruitment as primary RNAi effectors and use of small RNAs of certain preferred lengths. Prokaryotic PIWI proteins are typically components of several RNA-directed DNA restriction or CRISPR/Cas systems. However, eukaryotic versions appear to have emerged from a subset that evolved RNA-directed RNAi. They were recruited alongside RNaseIII domains and RNA-dependent RNA polymerase (RdRP) domains, also from prokaryotic systems, to form the core eukaryotic RNAi system. Like certain regulatory systems, RNAi diversified into two distinct but linked arms concomitant with eukaryotic nucleocytoplasmic compartmentalization. Subsequent elaboration of RNAi proceeded via diversification of the core protein machinery through lineage-specific expansions and recruitment of new components from prokaryotes (nucleases and small RNA-modifying enzymes), allowing for diversification of associating small RNAs.
近期,深度测序技术已广泛应用于各类真核生物的小RNA研究,极大地增进了我们对小RNA广泛参与调控多种生物学过程的理解。我们回顾了目前已知的小RNA类别,并将它们置于RNA干扰(RNAi)蛋白机制重建的进化史背景中。这种综合分析表明,真核生物RNAi系统的早期版本可能利用了从三种类型前体加工而来的小RNA:(1)正义-反义转录产物,(2)基因组编码的、不完全互补的发夹序列,以及(3)更大的非编码RNA前体序列。PIWI蛋白的结构剖析以及最近新家族(包括中介体复合物的Med13)的发现表明,通过祖先单元的复制形成的具有N端结构域(在原核生物中也单独与内切核酸酶融合)的独特结构的出现,是它们被招募为主要RNAi效应器并使用特定优选长度小RNA的关键。原核生物的PIWI蛋白通常是几种RNA导向的DNA限制或CRISPR/Cas系统的组成部分。然而,真核生物版本似乎起源于进化出RNA导向RNAi 的一个子集。它们与同样来自原核生物系统的RNaseIII结构域和RNA依赖性RNA聚合酶(RdRP)结构域一起被招募,形成了核心真核生物RNAi系统。与某些调节系统一样,RNAi随着真核生物核质区室化而多样化为两个不同但相互关联的分支。随后,RNAi通过核心蛋白机制的多样化得以进一步发展,这种多样化是通过谱系特异性扩展以及从原核生物中招募新成分(核酸酶和小RNA修饰酶)实现的,从而使得与之相关的小RNA得以多样化。