Molecular Biology Program, Memorial Sloan-Kettering Cancer Center, New York, New York 10021, USA;
Genes Dev. 2014 Mar 1;28(5):451-62. doi: 10.1101/gad.236745.113. Epub 2014 Feb 14.
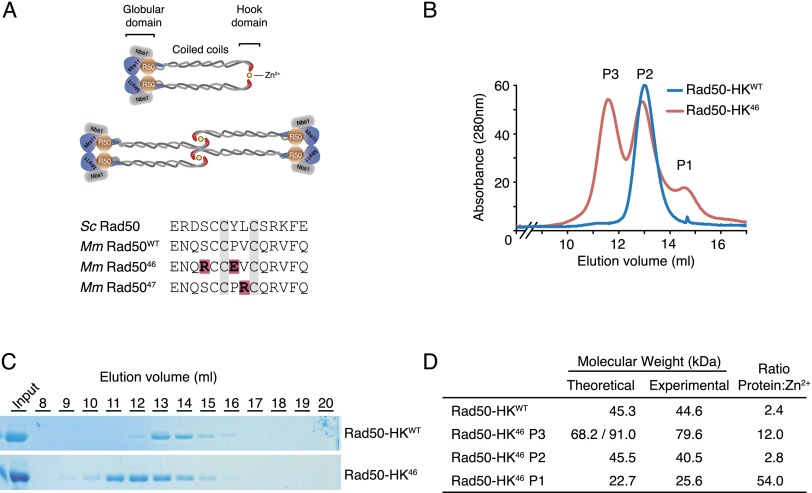
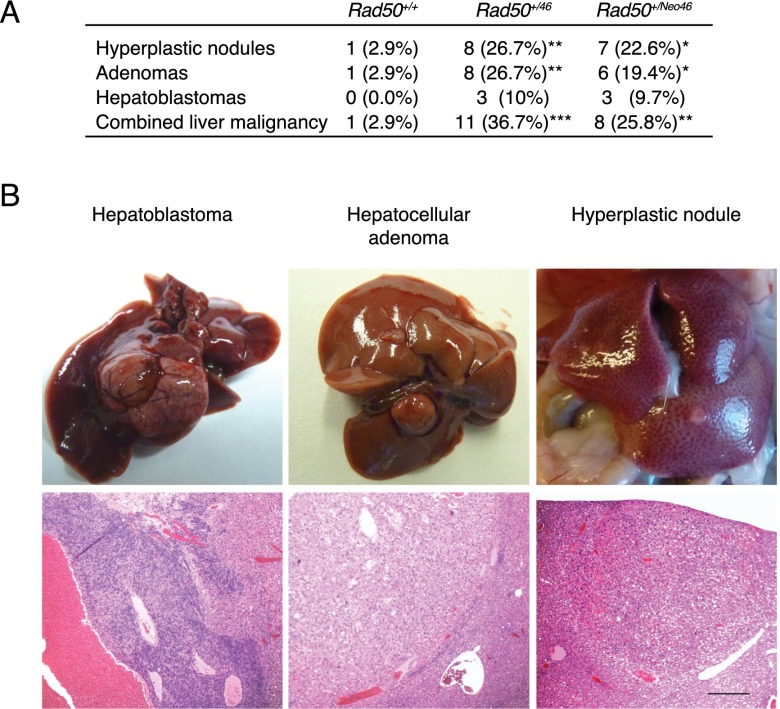
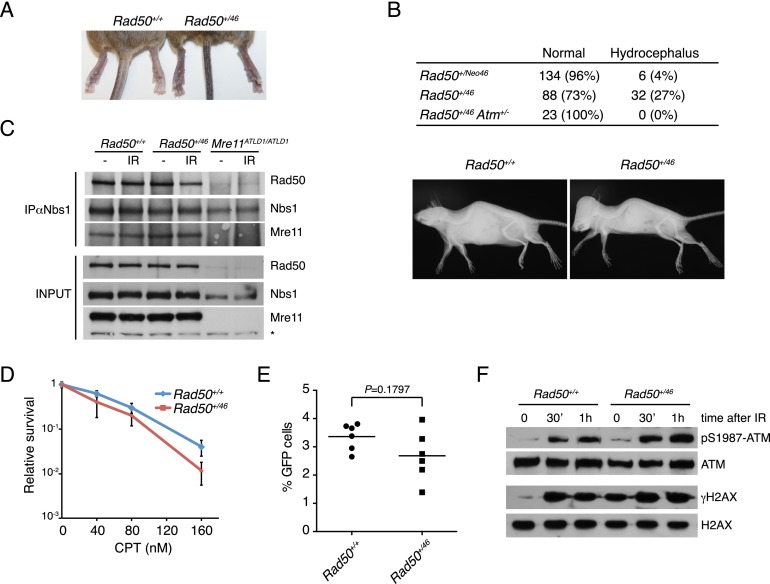
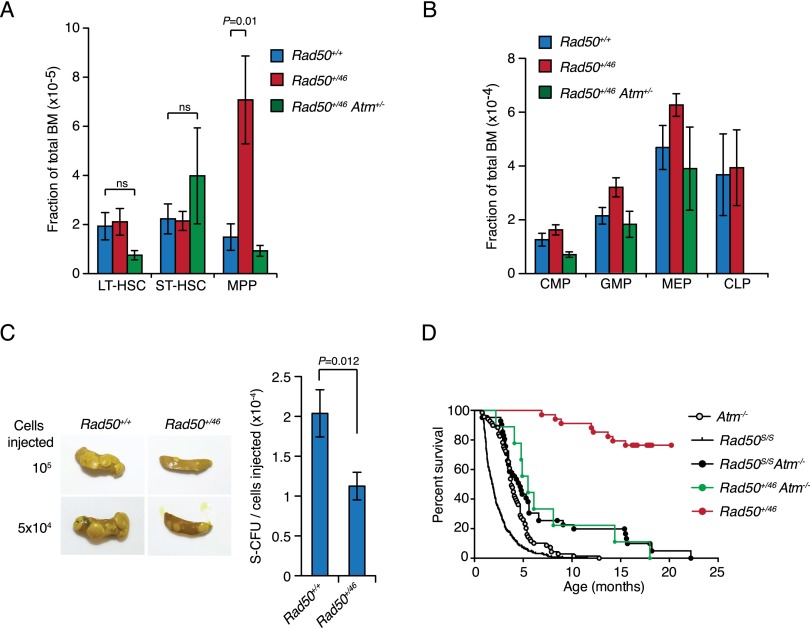
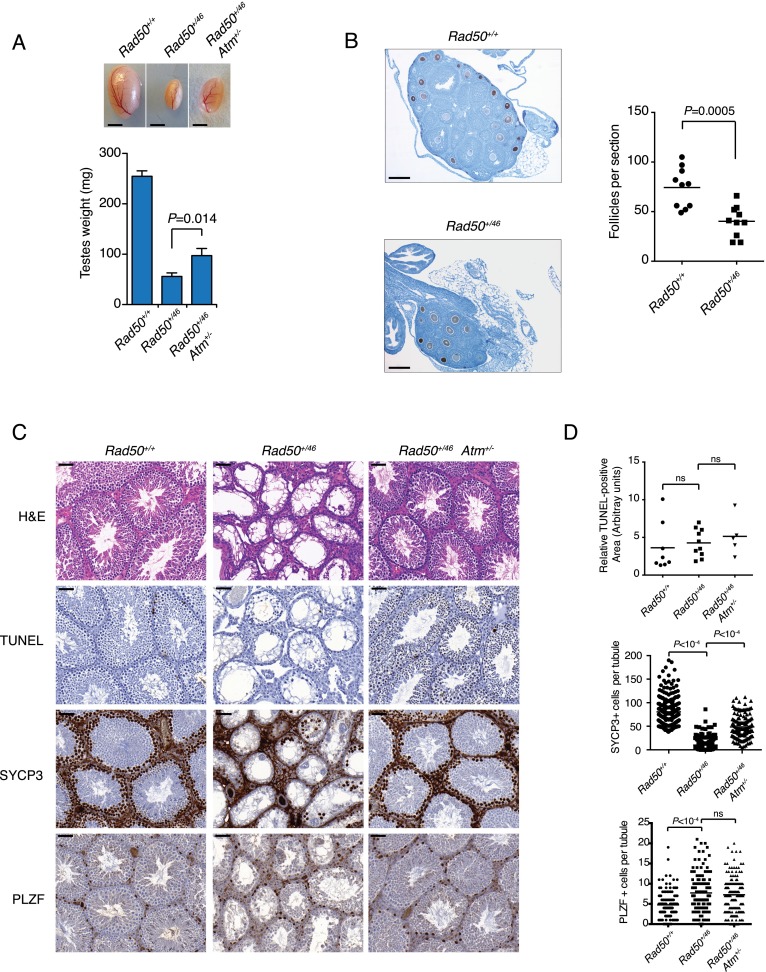
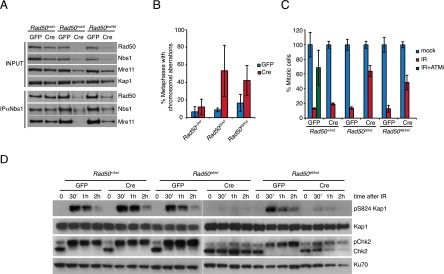
The Mre11 complex (Mre11, Rad50, and Nbs1) is a central component of the DNA damage response (DDR), governing both double-strand break repair and DDR signaling. Rad50 contains a highly conserved Zn(2+)-dependent homodimerization interface, the Rad50 hook domain. Mutations that inactivate the hook domain produce a null phenotype. In this study, we analyzed mutants with reduced hook domain function in an effort to stratify hook-dependent Mre11 complex functions. One of these alleles, Rad50(46), conferred reduced Zn(2+) affinity and dimerization efficiency. Homozygous Rad50(46/46) mutations were lethal in mice. However, in the presence of wild-type Rad50, Rad50(46) exerted a dominant gain-of-function phenotype associated with chronic DDR signaling. At the organismal level, Rad50(+/46) exhibited hydrocephalus, liver tumorigenesis, and defects in primitive hematopoietic and gametogenic cells. These outcomes were dependent on ATM, as all phenotypes were mitigated in Rad50(+/46) Atm(+/-) mice. These data reveal that the murine Rad50 hook domain strongly influences Mre11 complex-dependent DDR signaling, tissue homeostasis, and tumorigenesis.
Mre11 复合物(Mre11、Rad50 和 Nbs1)是 DNA 损伤反应(DDR)的核心组成部分,负责双链断裂修复和 DDR 信号转导。Rad50 包含一个高度保守的 Zn(2+)-依赖性同源二聚化界面,即 Rad50 钩结构域。失活钩结构域的突变会产生无效表型。在这项研究中,我们分析了具有降低的钩结构域功能的突变体,以努力分层钩依赖性 Mre11 复合物功能。这些等位基因之一,Rad50(46),赋予降低的 Zn(2+)亲和力和二聚化效率。纯合 Rad50(46/46)突变在小鼠中是致死的。然而,在野生型 Rad50 的存在下,Rad50(46)表现出与慢性 DDR 信号相关的显性获得性功能表型。在机体水平上,Rad50(+/46)表现出脑积水、肝肿瘤发生以及原始造血细胞和配子发生细胞的缺陷。这些结果依赖于 ATM,因为 Rad50(+/46) Atm(+/-) 小鼠减轻了所有表型。这些数据表明,鼠类 Rad50 钩结构域强烈影响 Mre11 复合物依赖性 DDR 信号转导、组织稳态和肿瘤发生。