European Molecular Biology Laboratory (EMBL), Mouse Biology Unit, Via Ramarini 32, Monterotondo Scalo 00015, Italy.
Genetics Department, Genomics Institute of the Novartis Research Foundation, 10675 John Jay Hopkins Drive, San Diego, CA 92121, USA.
Epigenetics Chromatin. 2014 Sep 11;7:24. doi: 10.1186/1756-8935-7-24. eCollection 2014.
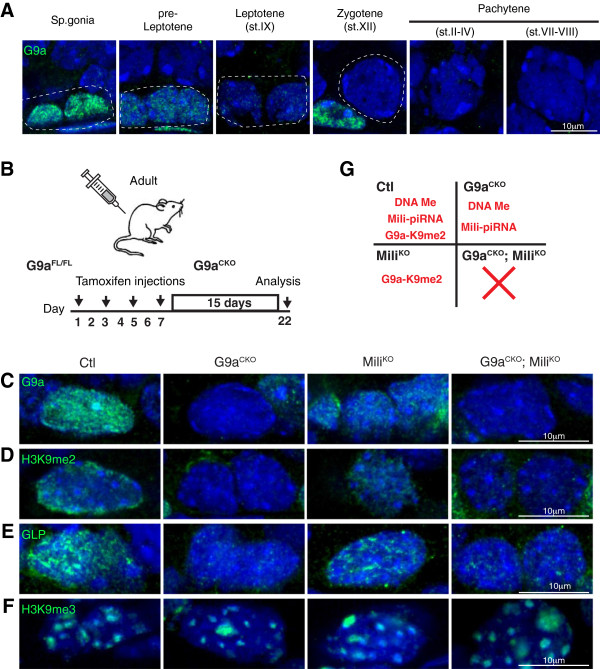
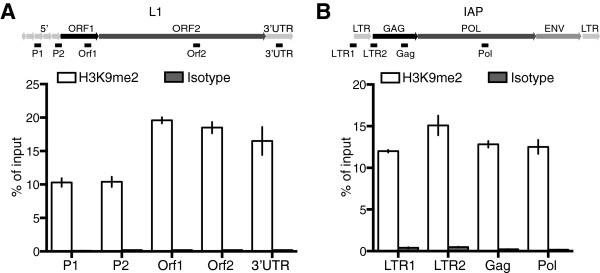
Repression of retrotransposons is essential for genome integrity and the development of germ cells. Among retrotransposons, the establishment of CpG DNA methylation and epigenetic silencing of LINE1 (L1) elements and the intracisternal A particle (IAP) endogenous retrovirus (ERV) is dependent upon the piRNA pathway during embryonic germ cell reprogramming. Furthermore, the Piwi protein Mili, guided by piRNAs, cleaves expressed L1 transcripts to post-transcriptionally enforce L1 silencing in meiotic cells. The loss of both DNA methylation and the Mili piRNA pathway does not affect L1 silencing in the mitotic spermatogonia where histone H3 lysine 9 dimethylation (H3K9me2) is postulated to co-repress these elements.
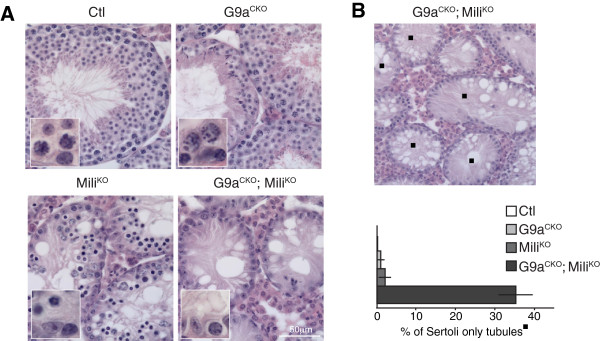
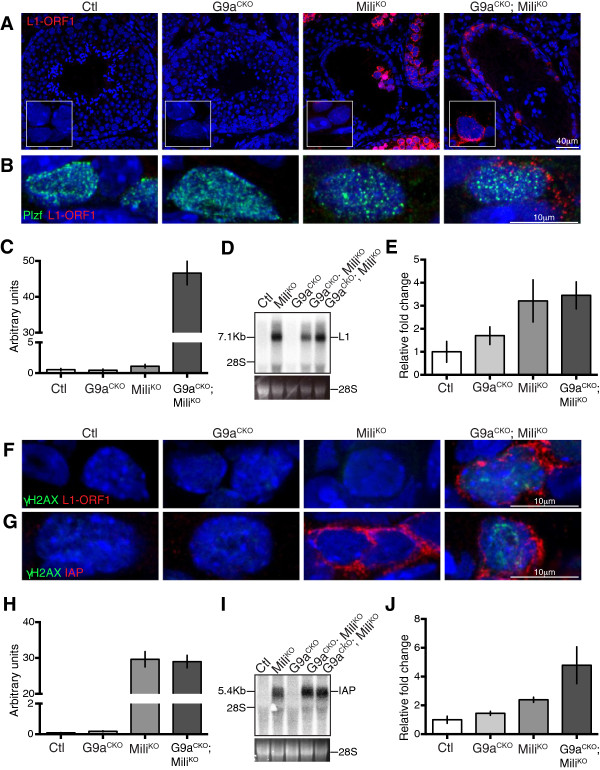
Here we show that the histone H3 lysine 9 dimethyltransferase G9a co-suppresses L1 elements in spermatogonia. In the absence of both a functional piRNA pathway and L1 DNA methylation, G9a is both essential and sufficient to silence L1 elements. In contrast, H3K9me2 alone is insufficient to maintain IAP silencing in spermatogonia. The loss of all three repressive mechanisms has a major impact on spermatogonial populations inclusive of spermatogonial stem cells, with the loss of all germ cells observed in a high portion of seminiferous tubules.
Our study identifies G9a-mediated H3K9me2 as a novel and important L1 repressive mechanism in the germ line. We also demonstrate fundamental differences in the requirements for the maintenance of L1 and IAP silencing during adult spermatogenesis, where H3K9me2 is sufficient to maintain L1 but not IAP silencing. Finally, we demonstrate that repression of retrotransposon activation in spermatogonia is important for the survival of this population and testicular homeostasis.
逆转录转座子的抑制对于基因组完整性和生殖细胞的发育至关重要。在逆转录转座子中,LINE1(L1)元件的 CpG DNA 甲基化和表观遗传沉默以及内源性逆转录病毒(ERV)的内膜 A 颗粒(IAP)的建立依赖于胚胎生殖细胞重编程过程中的 piRNA 途径。此外,Piwi 蛋白 Mili 指导 piRNA 切割表达的 L1 转录本,在后转录水平上在减数分裂细胞中强制执行 L1 沉默。DNA 甲基化和 Mili piRNA 途径的丧失都不会影响有丝分裂精原细胞中的 L1 沉默,据推测,组蛋白 H3 赖氨酸 9 二甲基化(H3K9me2)与共同抑制这些元件。
在这里,我们表明组蛋白 H3 赖氨酸 9 二甲基转移酶 G9a 共同抑制精原细胞中的 L1 元件。在缺乏功能性 piRNA 途径和 L1 DNA 甲基化的情况下,G9a 是沉默 L1 元件的必要和充分条件。相比之下,单独的 H3K9me2 不足以维持精原细胞中的 IAP 沉默。所有三种抑制机制的丧失对包括精原干细胞在内的精原细胞群体都有重大影响,在很大一部分生精小管中观察到所有生殖细胞的丧失。
我们的研究确定了 G9a 介导的 H3K9me2 是生殖系中 L1 的一种新的重要抑制机制。我们还证明了在成年精子发生过程中维持 L1 和 IAP 沉默的要求存在根本差异,其中 H3K9me2 足以维持 L1 沉默,但不能维持 IAP 沉默。最后,我们证明了精原细胞中逆转录转座子激活的抑制对于该细胞群体的存活和睾丸内稳态很重要。