National Leading Research Laboratory (NLRL) of Molecular Modeling and Drug Design, College of Pharmacy, Graduate School of Pharmaceutical Sciences, and Global Top 5 Research Program, Ewha Womans University, Seoul 120-750, Korea.
School of Computational Sciences, Korea Institute for Advanced Study, Seoul 130-722, Korea.
PLoS Comput Biol. 2015 Feb 9;11(2):e1004044. doi: 10.1371/journal.pcbi.1004044. eCollection 2015 Feb.
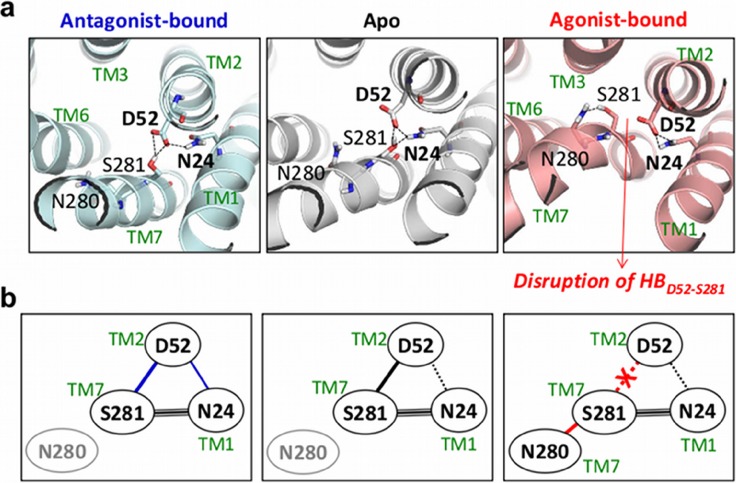
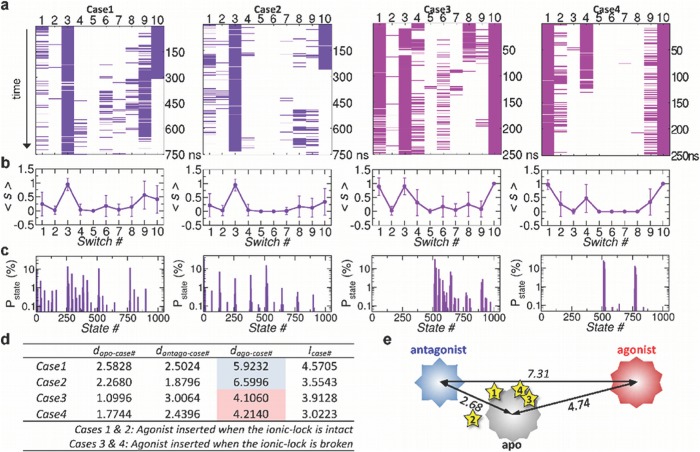
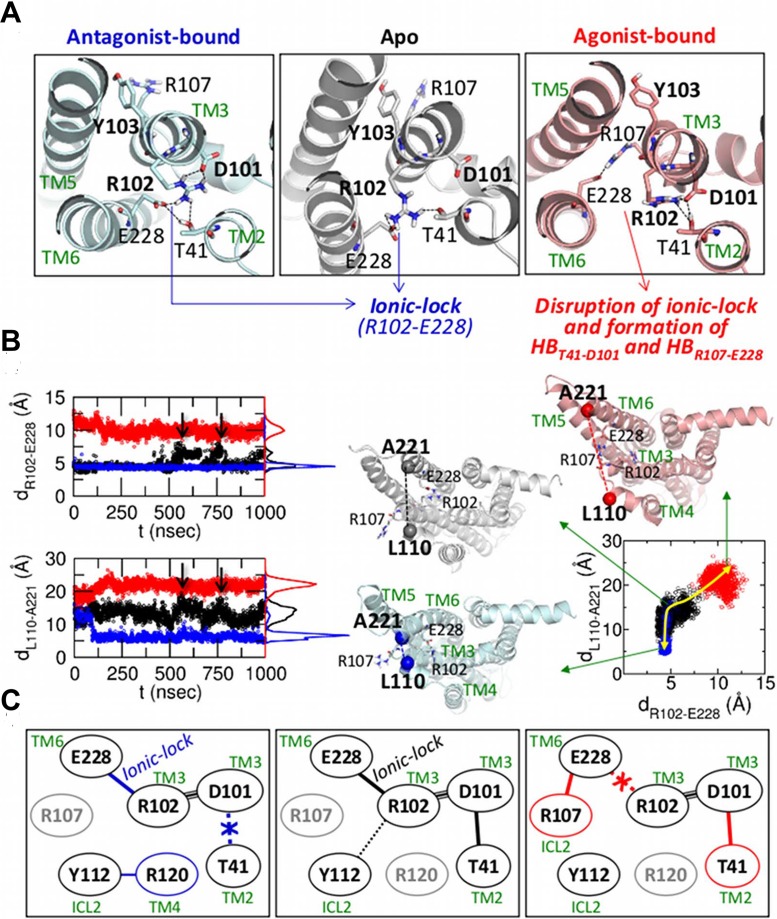
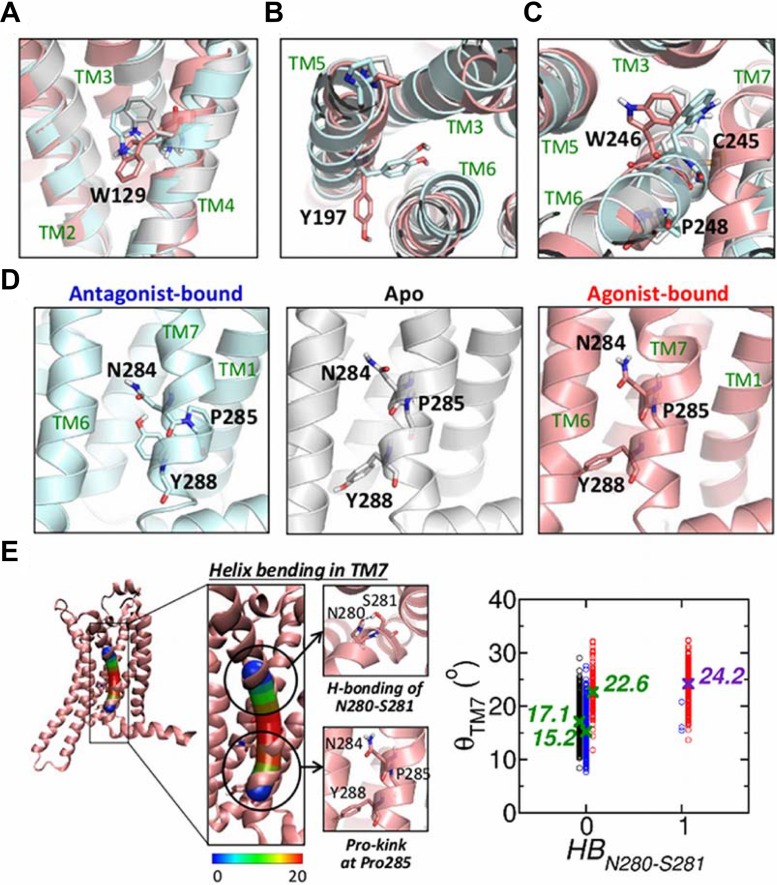
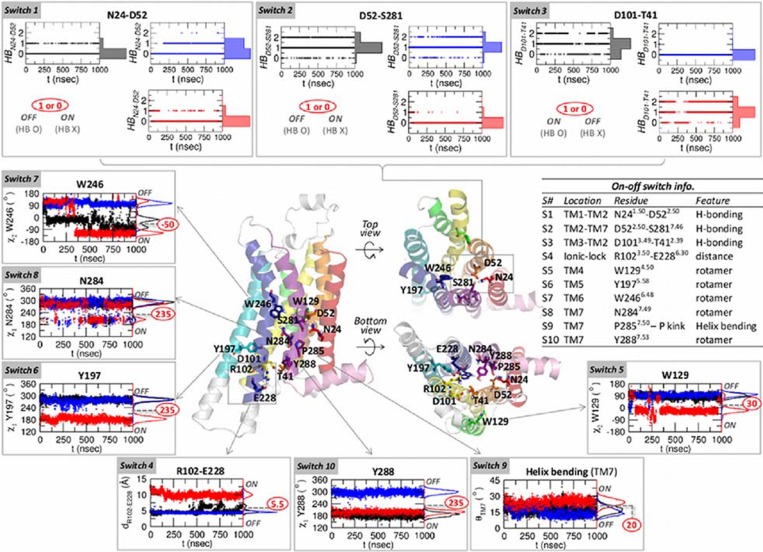
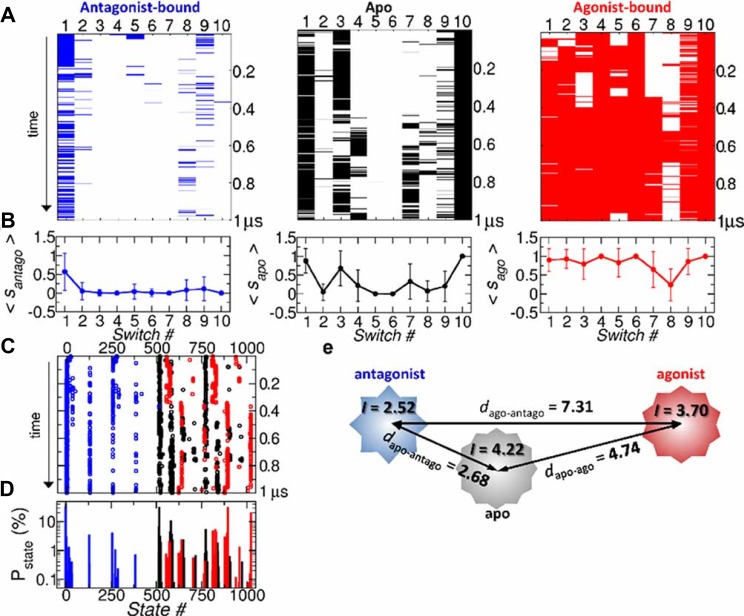
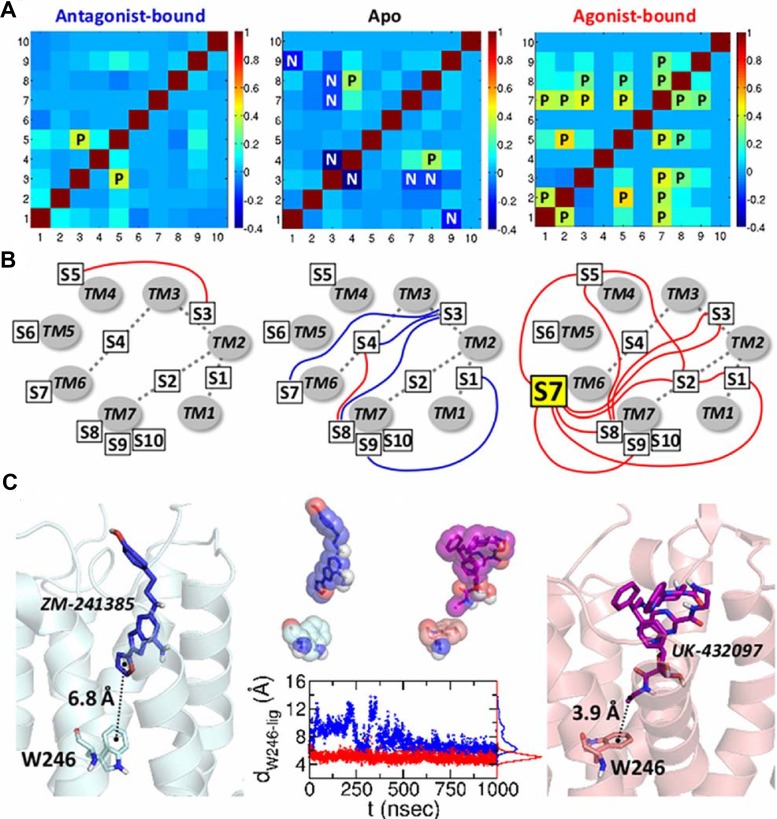
Dynamics and functions of G-protein coupled receptors (GPCRs) are accurately regulated by the type of ligands that bind to the orthosteric or allosteric binding sites. To glean the structural and dynamical origin of ligand-dependent modulation of GPCR activity, we performed total ~ 5 μsec molecular dynamics simulations of A2A adenosine receptor (A2AAR) in its apo, antagonist-bound, and agonist-bound forms in an explicit water and membrane environment, and examined the corresponding dynamics and correlation between the 10 key structural motifs that serve as the allosteric hotspots in intramolecular signaling network. We dubbed these 10 structural motifs "binary switches" as they display molecular interactions that switch between two distinct states. By projecting the receptor dynamics on these binary switches that yield 2(10) microstates, we show that (i) the receptors in apo, antagonist-bound, and agonist-bound states explore vastly different conformational space; (ii) among the three receptor states the apo state explores the broadest range of microstates; (iii) in the presence of the agonist, the active conformation is maintained through coherent couplings among the binary switches; and (iv) to be most specific, our analysis shows that W246, located deep inside the binding cleft, can serve as both an agonist sensor and actuator of ensuing intramolecular signaling for the receptor activation. Finally, our analysis of multiple trajectories generated by inserting an agonist to the apo state underscores that the transition of the receptor from inactive to active form requires the disruption of ionic-lock in the DRY motif.
G 蛋白偶联受体 (GPCRs) 的动力学和功能受到与其变构或正位结合位点结合的配体类型的精确调节。为了深入了解配体依赖性 GPCR 活性调节的结构和动力学起源,我们在明确的水相和膜环境中对 A2A 腺苷受体 (A2AAR) 的apo、拮抗剂结合和激动剂结合形式进行了总计约 5μsec 的分子动力学模拟,并检查了作为分子内信号网络变构热点的 10 个关键结构基序之间的相应动力学和相关性。我们将这 10 个结构基序称为“二进制开关”,因为它们显示出在两种不同状态之间切换的分子相互作用。通过将受体动力学投影到这些产生 2(10)微状态的二进制开关上,我们表明:(i) apo、拮抗剂结合和激动剂结合状态下的受体探索了截然不同的构象空间;(ii) 在这三种受体状态中,apo 状态探索的微状态范围最广;(iii) 在激动剂存在下,通过二进制开关之间的相干耦合维持了活性构象;(iv) 为了更具特异性,我们的分析表明,位于结合裂隙深处的 W246 可以作为激动剂传感器和引发受体激活的后续分子内信号的作用器。最后,我们对将激动剂插入 apo 状态生成的多个轨迹的分析强调了受体从不活跃形式向活跃形式的转变需要 DRY 基序中离子锁定的破坏。