Papp Stephanie J, Huber Anne-Laure, Jordan Sabine D, Kriebs Anna, Nguyen Madelena, Moresco James J, Yates John R, Lamia Katja A
Department of Chemical Physiology, Scripps Research Institute, La Jolla, United States.
Elife. 2015 Mar 10;4:e04883. doi: 10.7554/eLife.04883.
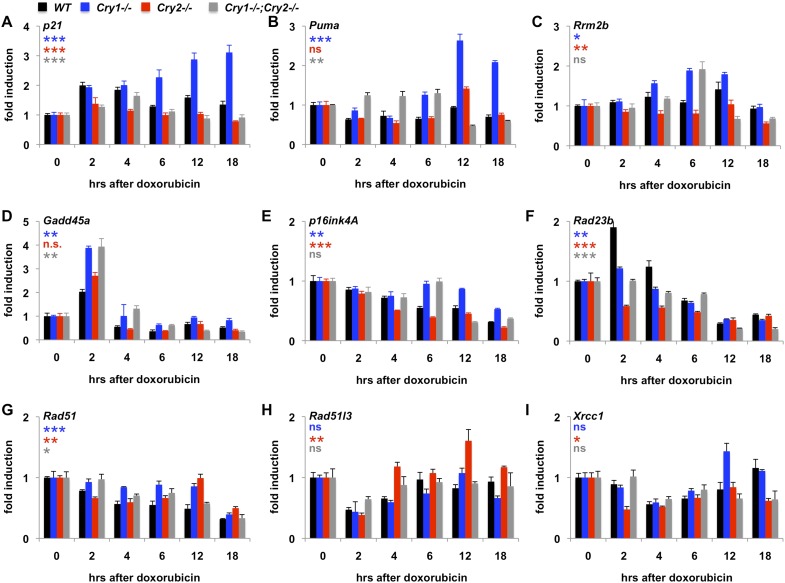
The circadian transcriptional repressors cryptochrome 1 (Cry1) and 2 (Cry2) evolved from photolyases, bacterial light-activated DNA repair enzymes. In this study, we report that while they have lost DNA repair activity, Cry1/2 adapted to protect genomic integrity by responding to DNA damage through posttranslational modification and coordinating the downstream transcriptional response. We demonstrate that genotoxic stress stimulates Cry1 phosphorylation and its deubiquitination by Herpes virus associated ubiquitin-specific protease (Hausp, a.k.a Usp7), stabilizing Cry1 and shifting circadian clock time. DNA damage also increases Cry2 interaction with Fbxl3, destabilizing Cry2. Thus, genotoxic stress increases the Cry1/Cry2 ratio, suggesting distinct functions for Cry1 and Cry2 following DNA damage. Indeed, the transcriptional response to genotoxic stress is enhanced in Cry1-/- and blunted in Cry2-/- cells. Furthermore, Cry2-/- cells accumulate damaged DNA. These results suggest that Cry1 and Cry2, which evolved from DNA repair enzymes, protect genomic integrity via coordinated transcriptional regulation.
昼夜节律转录抑制因子隐花色素1(Cry1)和隐花色素2(Cry2)由光解酶进化而来,光解酶是细菌中的光激活DNA修复酶。在本研究中,我们报告称,虽然Cry1/2已丧失DNA修复活性,但它们通过翻译后修饰响应DNA损伤并协调下游转录反应,从而适应保护基因组完整性。我们证明,遗传毒性应激刺激Cry1磷酸化及其被疱疹病毒相关泛素特异性蛋白酶(Hausp,又名Usp7)去泛素化,使Cry1稳定并改变昼夜节律时钟时间。DNA损伤还会增加Cry2与Fbxl3的相互作用,使Cry2不稳定。因此,遗传毒性应激会增加Cry1/Cry2的比例,表明DNA损伤后Cry1和Cry2具有不同的功能。事实上,对遗传毒性应激的转录反应在Cry1-/-细胞中增强,而在Cry2-/-细胞中减弱。此外,Cry2-/-细胞会积累受损DNA。这些结果表明,从DNA修复酶进化而来的Cry1和Cry2通过协调转录调控来保护基因组完整性。